Der Entzündungsfaktor, der den meisten Krebsarten zugrunde liegt
 Der Beginn von 95% aller Krebserkrankungen ist ein weit verbreitetes, aber wenig bekanntes Molekül, von dem die meisten Menschen noch nie etwas gehört haben: Nuklearfaktor-kappaB oder NF-kB.1-3 Neue Forschung bestätigt die Gefahr, die sie für erwachsene Menschen darstellt.
Der Beginn von 95% aller Krebserkrankungen ist ein weit verbreitetes, aber wenig bekanntes Molekül, von dem die meisten Menschen noch nie etwas gehört haben: Nuklearfaktor-kappaB oder NF-kB.1-3 Neue Forschung bestätigt die Gefahr, die sie für erwachsene Menschen darstellt.
Zum Beispiel hat eine Ende des Jahres veröffentlichte wegweisende Studie4 zum ersten Mal die zentrale Rolle von NF-kB bei der Entstehung von malignem Brustkrebs bestätigt.
Ein Forscherteam fand heraus, dass Brustkrebszellen eine spezifische Entzündungskaskade benötigen, um sich zu vermehren und zu metastasieren.
NF-kB ist eines der Proteine, die an der Entzündung beteiligte Gene aktivieren.
Glücklicherweise gibt es viele Nährstoffe, die die Mitglieder von Life Extension® bereits einnehmen, von dem gezeigt wurde, dass die krebsverursachende Entzündungskaskade, vermittelt durch NF-kB, sich neutralisiert und sogar umgekehrt .
In diesem Artikel wird eine umfassende Übersicht über NF-kB-inhibierende Verbindungen beschrieben. Sie erfahren, wie NF-kB das Wachstum und die Entwicklung vieler Arten von Tumoren auslöst und als Hauptschalter für Zellen dient, die mit krebserregenden Genen "vorgeladen" sind. Sie werden aussagekräftige Beweise für kostengünstige Interventionen finden, die NF-kB unterdrücken, tödliche Krebsgene ausschalten und die Entzündungskaskade, die Krebszellen zur Ausbreitung benötigen, unterbrechen.
Seit seiner Entdeckung im Jahr 1986 ist bekannt, dass NF-kB eng mit der Entwicklung von Krebs in Verbindung steht.5 Heute wissen wir, dass die meisten, wenn nicht alle Krebsarten ungewöhnlich hohe Mengen an aktivem NF-kB aufweisen, was sie in einem anhaltenden Entzündung Zustand hält.6 Umgekehrt wirken bestimmte sogenannte Onkogene (abnormale Gene, die eine Krebsentwicklung auslösen können), indem sie NF-kB aktivieren. Zusammen bilden diese Tatsachen einen konstanten Strom entzündlicher und krebsfördernder Cytokine innerhalb eines wachsenden Tumors.8
Die Produktion dieser Zellsignalmoleküle macht es sehr viel wahrscheinlicher, dass eine bestimmte krebsanfällige Zelle einen bösartigen Tumor bildet. Dies kann auch bedeuten, dass ein bestimmter Tumor gegenüber Bestrahlung und Chemotherapie viel resistenter ist, da NF-kB im Allgemeinen das Überleben von Krebszellen fördert.9
Die NF-kB-Krebsblockade
 NF-kB löst eine Kaskade von Ereignissen aus, die eine Entzündung mit der Krebsentwicklung verbinden. Sobald dieser Prozess eingeleitet wurde, feuert NF-kB die Flammen weiter an und trägt so zur Invasivität der Krebszellen, zur Stimulierung einer frischen Blutversorgung und zur Ausbreitung durch Metastasen im gesamten Körper bei.
NF-kB löst eine Kaskade von Ereignissen aus, die eine Entzündung mit der Krebsentwicklung verbinden. Sobald dieser Prozess eingeleitet wurde, feuert NF-kB die Flammen weiter an und trägt so zur Invasivität der Krebszellen, zur Stimulierung einer frischen Blutversorgung und zur Ausbreitung durch Metastasen im gesamten Körper bei.
Aber unser Wissen über den Schaden, den NF-kB angerichtet hat, kann zum Guten wenden. Sobald er entdeckt wurde, erkannten vorausschauende Forscher, dass sie die Aktivierung von NF-kB blockieren könnten.
Wir wissen beispielsweise seit fast 12 Jahren, dass wir Entzündungen in Darmkrebszellen durch Behandlung mit dem Gewürz Curcumin, einem bekannten Inhibitor der NF-kB-Aktivierung, reduzieren können.10 Die Blockierung der NF-kB-Aktivierung kann auch die menschliche Prostata Krebszellen zur Apoptose sensibilisieren - der programmierte Zelltod, der Krebserkrankungen unter Kontrolle hält.11 Und natürliche Moleküle, die NF-kB hemmen, können entzündliche Veränderungen des Pankreas durch Alkohol einschränken.12
Das Potenzial zur Blockierung von NF-kB bietet Hoffnung im Zusammenhang mit dem tödlichen HER-2-Brustkrebsgen. HER-2-Krebserkrankungen sind normalerweise sehr therapieresistent und haben eine schlechte Prognose.6 Die Blockade von NF-kB kann die Brustkrebszellen für eine Chemotherapie besser zugänglich machen.8,13
Es ist nicht überraschend, dass die großen Pharmakonzerne intensiv daran interessiert sind, neue (und rentable) Medikamente zu entdecken, die NF-kB blockieren.14 Die Natur hat uns jedoch eine beträchtliche Anzahl von Substanzen geliefert, die die proinflammatorischen krebsbekämpfenden Wirkungen von Renegade NF -kB blockieren oder hemmen - und sie sind sofort verfügbar, erschwinglich und sicher. Schauen wir uns ein Beispiel an.
Nährstoffe für die entzündlichen krebsbekämpfenden Wirkungen von NF-kB
 Nach einigen Schätzungen könnten 67% bis 90% der Krebserkrankungen beim Menschen durch Modifikationen des Lebensstils, insbesondere der Ernährung, verhindert werden.14,15 Die Unterdrückung von NF-kB durch in Früchten, Gemüse und vielen Gewürzen enthaltene Phytochemikalien wurde die molekulare Basis für ihre Fähigkeit zur Krebsprävention zu schaffen bekannt.16,17
Nach einigen Schätzungen könnten 67% bis 90% der Krebserkrankungen beim Menschen durch Modifikationen des Lebensstils, insbesondere der Ernährung, verhindert werden.14,15 Die Unterdrückung von NF-kB durch in Früchten, Gemüse und vielen Gewürzen enthaltene Phytochemikalien wurde die molekulare Basis für ihre Fähigkeit zur Krebsprävention zu schaffen bekannt.16,17
Die folgenden Nutrazeutika können uns der erfolgreichen Chemoprävention näher bringen, indem sie die proinflammatorischen Wirkungen von NF-kB gegen Krebs einsetzen können. Im Gegensatz zu Medikamenten mit Einzelzielen sind die meisten Nutrazeutika mehrzielend. Dies verleiht ihnen erstaunliche Effizienz und kann ihre Wirksamkeit steigern, insbesondere wenn komplexe Signalwege wie bei NF-kB18 unterbrochen werden
Antioxidantien
 Die oxidative Schädigung von DNA, Proteinen und anderen lebenswichtigen Strukturen ist ein wesentlicher Faktor für die Entstehung, Förderung und Verbreitung von Krebserkrankungen. Dies macht Antioxidantien zu wichtigen Mitteln bei der Bekämpfung von Krebs.19 Viele Antioxidantien können jedoch auch NF-kB blockieren, wodurch deren Wirkungen eine wertvolle Rolle spielen. Sie tun dies über eine Reihe eng miteinander verbundener, aber unterschiedlicher Mechanismen.14
Die oxidative Schädigung von DNA, Proteinen und anderen lebenswichtigen Strukturen ist ein wesentlicher Faktor für die Entstehung, Förderung und Verbreitung von Krebserkrankungen. Dies macht Antioxidantien zu wichtigen Mitteln bei der Bekämpfung von Krebs.19 Viele Antioxidantien können jedoch auch NF-kB blockieren, wodurch deren Wirkungen eine wertvolle Rolle spielen. Sie tun dies über eine Reihe eng miteinander verbundener, aber unterschiedlicher Mechanismen.14
Ein wichtiger Weg, die NF-kB-Aktivität zu blockieren, besteht darin, zu verhindern, dass sie durch den Eintritt in den Zellkern aktiviert wird. Dies verweigert NF-kB den Zugang zu den proinflammatorischen Genen, die es reguliert, und verhindert so die Entzündungsreaktion, bevor sie beginnt. Zellen mit größeren Antioxidationsmittel-Abwehrkräften haben im Allgemeinen eine geringere NF-kB-Aktivierung. Die Erhöhung dieser Abwehrkräfte korreliert mit einer reduzierten NF-kB-Expression in Zellkernen.20,21
Eine große Anzahl von Antioxidantien wird hinsichtlich spezifischer NF-kB-Blockierungsaktivität geprüft. Dazu gehören die Vitamine C und E, Carotinoide, die sogenannten Thiol-Antioxidantien (Glutathion, Thioredoxin und Liponsäure), Flavonoide und Polyphenole, Selen, Zink und andere.14,19, 22, 23 Das Flavonoid Luteolin kann beispielsweise Krebszellen für Chemotherapeutika sensibilisieren, indem es verschiedene Zellüberlebenswege unterdrückt, darunter NF-kB. Zink, ein essentielles Spurenelement, hemmt NF-kB indirekt durch Induzieren eines separaten genannten regulatorischen Proteins A.20.25
 Mariendistelextrakte verbessern den Zelltod des Prostatakrebses durch Apoptose, indem sie den NF-kB-Spiegel im Zellkern senken.26,27 Das Tomatencarotinoid-Pigment Lycopin verhindert die Aktivierung von NF-kB beim Prostatakrebs durch einen anderen Mechanismus.28 Eine Form von Vitamin E, genannt Gamma-Tocotrienol, senkt NF-kB in Tumorzellen der Bauchspeicheldrüse.29 Selbst bestimmte Kaffeekomponenten, die als Diterpene bezeichneten Antioxidationsmoleküle, unterdrücken die Aktivierung von NF-kB.31 Schließlich reduziert das Tomatenpolyphenol-Lycopin die NF-kB-Aktivierung in einer Vielzahl von Krebszelltypen.28
Mariendistelextrakte verbessern den Zelltod des Prostatakrebses durch Apoptose, indem sie den NF-kB-Spiegel im Zellkern senken.26,27 Das Tomatencarotinoid-Pigment Lycopin verhindert die Aktivierung von NF-kB beim Prostatakrebs durch einen anderen Mechanismus.28 Eine Form von Vitamin E, genannt Gamma-Tocotrienol, senkt NF-kB in Tumorzellen der Bauchspeicheldrüse.29 Selbst bestimmte Kaffeekomponenten, die als Diterpene bezeichneten Antioxidationsmoleküle, unterdrücken die Aktivierung von NF-kB.31 Schließlich reduziert das Tomatenpolyphenol-Lycopin die NF-kB-Aktivierung in einer Vielzahl von Krebszelltypen.28
Die Liste der antioxidativen Nährstoffe mit NF-kB-unterdrückender Wirkung wächst deutlich. Eine gute allgemeine Regel für die Gegenwart besteht darin, sicherzustellen, dass Sie eine ausreichende Menge an wirksamen, hochwertigen Antioxidantien als Teil Ihres Gesundheitsplans einbeziehen. Wenden wir uns nun einigen spezifischen Nährstoffen zu, die zusätzliche NF-kB-Blocking-Effekte haben, die Ihren Krebsschutz optimieren können.
- Eine Durchbruchstudie ergab, dass eine Entzündung notwendig ist, damit sich Brustkrebszellen vermehren und metastasieren können.
- Der Master Control-Komplex namens Nuclear Factor-kappaB oder NF-kB löst und reguliert diese Entzündungskaskade, die mit dem Beginn und der Entwicklung von 95% aller Krebserkrankungen in Verbindung steht.
- NF-kB stimuliert Gene zur Produktion entzündlicher Zytokine und anderer Signalmoleküle, die das Wachstum und die Entwicklung von Krebs fördern.
- Die Hemmung von NF-kB ist ein wirksames Ziel für die Krebsprävention und es wurde gezeigt, dass es krebsstimulierende Ereignisse auf allen Ebenen der Karzinogenese (Krebsentwicklung) verringert.
- Eine breite Palette natürlicher diätetischer Interventionen hemmt die pathologische Aktivität von NF-kB auf kraftvolle Weise. Dies ist eine wichtige Strategie im Kampf gegen Krebs.
Curcumin
 Curcumin ist ein außergewöhnliches Molekül, das aus dem Gewürz gewonnen wird, das wir als Kurkuma kennen. Es gehört zu den stärksten der natürlichen Inhibitoren von NF-kB und wurde daher hinsichtlich seines Antikrebspotenzials intensiv untersucht.32
Curcumin ist ein außergewöhnliches Molekül, das aus dem Gewürz gewonnen wird, das wir als Kurkuma kennen. Es gehört zu den stärksten der natürlichen Inhibitoren von NF-kB und wurde daher hinsichtlich seines Antikrebspotenzials intensiv untersucht.32
Curcumin ist bei allen drei Hauptstadien der Karzinogenese von Nutzen: Einleitung, Fortschreiten und Beförderung.33 Der größte Teil dieses Nutzens steht im Zusammenhang mit seiner Hemmung von NF-kB und der nachfolgenden Hemmung der proinflammatorischen Signalgebung.33 Die NF-kB-Hemmung wird ebenfalls zugeschrieben mit der Fähigkeit von Curcumin, die Proliferation von Krebszellen, die Invasion, die Bildung neuer Blutgefäße und die Metastasierung zu hemmen.34 Eine weitere wichtige Folge der NF-kB-Hemmung durch Curcumin besteht darin, den natürlichen Verlauf der Apoptose in Krebszellen ablaufen zu lassen, so dass sie absterben anstatt sich zu vermehren und zu verbreiten.35
Curcumin ist vielversprechend, wenn es darum geht, einige der häufigsten und gefährlichsten Malignome der Menschheit zu behandeln oder zu verhindern. Dazu gehören chronische lymphatische Leukämien, Lymphome, Bauchspeicheldrüsenkrebs, Blasenkrebs, Dickdarmkrebs und Gallengangskrebs.36-44 Curcumin kann dazu beitragen, die durch Fettleibigkeit hervorgerufene krebserregende Entzündung, eines der dringlichsten Gesundheitsprobleme der Gegenwart, zu unterdrücken. 45
NF-kB ist wissenschaftlich als "Transkriptionsfaktor" bekannt. 92 Es regelt, wie und wann die Informationen in bestimmten menschlichen Genen in neue Proteinmoleküle übersetzt werden.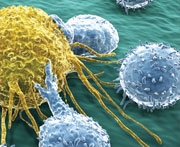
Zu diesen Genen zählen vor allem solche, die Entzündungsreaktionen fördern, von denen viele auch das Zellwachstum, das Überleben und die Umwandlung in Krebs fördern. Tatsächlich wurde Anfang der 2000er Jahre klar, dass NF-kB das „fehlende Bindeglied“ zwischen den beiden, Entzündung und Krebs, ist.93 Natürlich ist das nicht die einzige Rolle.
Bei gesunden Menschen erfüllt NF-kB eine wichtige Funktion, indem es inaktiv bleibt. Dies hilft, die leistungsstarke zelluläre Maschinerie der Entzündung im Leerlauf zu halten. Wenn eine Bedrohung auftritt, wird NF-kB jedoch aktiviert und arbeitet innerhalb der Zellkerne (daher der Begriff „Kernfaktor“). 7
Im Zellkern werden alle genetischen Informationen in Form von DNA in unseren Chromosomen gespeichert. Unter normalen Umständen „lebt“ NF-kB nicht im Zellkern, sondern im Zytoplasma der Zelle, dem Hauptkörper der Zelle.7 Wenn eine Infektion oder ein anderer fremder Eindringling droht, wandert NF-kB aus dem Zytoplasma der Zelle in den Zellkern. 7 Sobald sich NF-kB im Zellkern befindet, bindet es an bestimmte Teile des Chromosoms, wo es die Transkription eines Gens in sein Proteinprodukt aktiviert.
Viele Gene, die unter die Kontrolle von NF-kB geraten, sind proinflammatorische Gene, das heißt, ihre Produkte sind entzündliche Cytokine wie Interleukine, Tumornekrosefaktor und andere. Sobald der Entzündungsreiz weg ist, löst sich NF-kB normalerweise vom Chromosom und kehrt zum Zytoplasma zurück, und das Entzündungsgen stellt die Produktion seiner Zytokine ein. Mit anderen Worten, unter normalen Umständen stirbt eine Entzündung ab, wenn sie nicht mehr von Nutzen ist.94
Aber das nur unter normalen Umständen und Krebs ist alles andere als normal.
Knoblauch-Extrakte
 Knoblauch hat wie Kurkuma in traditionellen medizinischen Systemen eine lange Tradition. Wenn Knoblauch-Extrakte altern gelassen werden, stabilisieren sie sich und behalten starke antioxidative Kräfte bei, die NF-kB indirekt herunterregulieren.46 Einzelne Bestandteile dieser Knoblauchextrakte umfassen Diallylsulfid, Diallyldisulfid und Thiacremonon, die alle direkte NF-kB- inhibitorische Aktivitäten.17,47,48
Knoblauch hat wie Kurkuma in traditionellen medizinischen Systemen eine lange Tradition. Wenn Knoblauch-Extrakte altern gelassen werden, stabilisieren sie sich und behalten starke antioxidative Kräfte bei, die NF-kB indirekt herunterregulieren.46 Einzelne Bestandteile dieser Knoblauchextrakte umfassen Diallylsulfid, Diallyldisulfid und Thiacremonon, die alle direkte NF-kB- inhibitorische Aktivitäten.17,47,48
Die Hemmung von NF-kB durch Knoblauchextrakte ist vielversprechend bei der Verhinderung des Wachstums und der Ausbreitung vieler Tumortypen, einschließlich Darmkrebs, malignem Neuroblastom und Melanom.47-51 Knoblauchextrakte induzieren auch den Zelltod von Krebs durch Apoptose.49,51
Ingwer
 Ingwer ist ein Gewürz aus Südasien, das viele gesundheitsfördernde Bestandteile enthält.52 Gingerol ist dessen Hauptstoff; Es ist eine phenolische Verbindung mit antioxidativen, entzündungshemmenden und tumorhemmenden Eigenschaften.53 Gingerol unterdrückt Entzündungen, indem es die Bewegung von NF-kB in den Zellkern blockiert, wodurch entzündliche Zytokine wie TNF-alpha herunterreguliert werden .52,53 Eine weitere wichtige Ingwer-Verbindung, Zerumbon, unterdrückt die durch eine Vielzahl von üblichen Karzinogenen induzierte NF-kB-Aktivierung, blockiert Metastasenbildung, Invasion und erhöht gleichzeitig die Apoptose.54
Ingwer ist ein Gewürz aus Südasien, das viele gesundheitsfördernde Bestandteile enthält.52 Gingerol ist dessen Hauptstoff; Es ist eine phenolische Verbindung mit antioxidativen, entzündungshemmenden und tumorhemmenden Eigenschaften.53 Gingerol unterdrückt Entzündungen, indem es die Bewegung von NF-kB in den Zellkern blockiert, wodurch entzündliche Zytokine wie TNF-alpha herunterreguliert werden .52,53 Eine weitere wichtige Ingwer-Verbindung, Zerumbon, unterdrückt die durch eine Vielzahl von üblichen Karzinogenen induzierte NF-kB-Aktivierung, blockiert Metastasenbildung, Invasion und erhöht gleichzeitig die Apoptose.54
Ingwer-Extrakte wurden verwendet, um die Lebensfähigkeit von Magenkrebszellen, Eierstockkrebs, Brust-, Leber-, Darm- und Lungenkrebs sowie UV-Licht-induzierte Hautkrebserkrankungen zu reduzieren.55-60
Dank Dr. Sahdeo Prasad vom Cytokine Research Laboratory des MD Anderson Cancer Center in Houston, wissen wir im Jahr 2011 über die Beziehung zwischen NF-kB und Krebs folgendes: 5
NF-kB wird als Reaktion auf eine Reihe von Faktoren aktiviert, die bis zu 95% aller Krebserkrankungen ausmachen. Diese Auslöser umfassen Tabak, Stress, diätetische Komponenten, Fettleibigkeit, Alkohol, Infektionen, Strahlung und Umweltgifte.1-3
Die Aktivierung von NF-kB wurde mit der Umwandlung normaler Zellen in Krebszellen in Verbindung gebracht (daher kann man davon ausgehen, dass NF-kB die Krebsentwicklung fördert) .1,95
NF-kB ist in Krebszellen mit einer höheren Rate als in gesunden Zellen aktiv. Ein sich entwickelnder Krebs erhöht auch die Menge an aktivem NF-kB.6,13
NF-kB wurde mit dem Überleben gefährlicher Krebsstammzellen in Verbindung gebracht, den frühen "Elternzellen" vieler Krebsarten, die heute in ansonsten normalen Geweben bekannt sind. Krebsstammzellen können sich erfolgreicher als gesunde Zellen "selbst erneuern".
NF-kB stimuliert die Produktion von Genprodukten, die verhindern, dass Krebszellen durch den Prozess des programmierten Zelltods oder durch Apoptose auf natürliche Weise absterben.96
NF-kB erhöht auch die Produktion von Genprodukten im Zusammenhang mit der Proliferation, der schnellen und wiederholten Zellteilung, die Krebsarten zu aggressiven Wachstumseigenschaften verhelfen.97
NF-kB kontrolliert die Expression von Genprodukten, die mit der Invasion, dem Wachstum neuer Blutgefäße (Angiogenese) und der metastatischen Ausbreitung von Tumoren zusammenhängen.96,98-100
Viele Karzinogene aktivieren NF-kB, aber wirksamste krebspräventive (chemopräventive) Verbindungen unterdrücken die Aktivierung von NF-kB.1,15,101
Prasad weist darauf hin, dass diese Beobachtungen zeigen, wie eng NF-kB mit Krebswachstum und Metastasierung verknüpft ist.5 Andere weisen darauf hin, dass NF-kB zwar ein mächtiger Nemesis ist, aber auch unzählige Möglichkeiten bietet, kraftvoll eingreifen zu können und Krebserkrankungen in ihren Spuren zu blockieren.8,13 Während wir die enge Beziehung zwischen chronischen Entzündungen und Krebs weiter klarstellen, werden immer mehr solche Gelegenheiten offensichtlich.93
Grüner Tee
 Extrakte aus grünem Tee sind weithin für ihre Vorteile bei der Prävention vieler häufiger chronischer Erkrankungen bekannt. Ihre Hauptbestandteile, die Catechinfamilie der Polyphenole, sind starke Antioxidantien, denen die meisten positiven Wirkungen des Tees zu verdanken sind.61,62 Catechine wirken auf viele verschiedene Ziele.18,63,64 Die Inhibierung von NF-kB durch Catechine des grünen Tees hat kürzlich stattgefunden. Es wurde festgestellt, dass dies ein Hauptmechanismus ist, durch den sie jede Stufe der Karzinogenese blockiert.63-65
Extrakte aus grünem Tee sind weithin für ihre Vorteile bei der Prävention vieler häufiger chronischer Erkrankungen bekannt. Ihre Hauptbestandteile, die Catechinfamilie der Polyphenole, sind starke Antioxidantien, denen die meisten positiven Wirkungen des Tees zu verdanken sind.61,62 Catechine wirken auf viele verschiedene Ziele.18,63,64 Die Inhibierung von NF-kB durch Catechine des grünen Tees hat kürzlich stattgefunden. Es wurde festgestellt, dass dies ein Hauptmechanismus ist, durch den sie jede Stufe der Karzinogenese blockiert.63-65
NF-kB-Inhibierung durch Grüntee-Extrakte, die reich an Epigallocatechingallat (EGCG) sind, ist von besonderem Interesse.66 Es wurde gezeigt, dass EGCG bei experimentellem Prostatakrebs Apoptose produziert, das Wachstum von Plattenepithelkarzinomen des Kopfes und Halses hemmt, die Produktion einschränkt und hemmt. Die Invasion von experimentell induzierten Brustkrebserkrankungen reduziert das Auftreten karzinogeninduzierter Lungenkrebserkrankungen und sensibilisiert Melanomzellen für die Wachstumshemmung durch andere Erreger.67-71
Isoflavone
 Isoflavone und Isoflavonoide sind Moleküle, die in vielen Pflanzen, insbesondere in der Bohnenfamilie, vorkommen. Pflanzen verwenden sie als Mittel zur Bekämpfung von Krankheiten und als Stimulanzien für die symbiotischen Bakterien, die ihnen helfen, Stickstoff zum Aufbau von Proteinen zu verwenden. Isoflavone aus Sojabohnen haben auch beim Menschen viele positive und schützende Wirkungen. Die niedrige Rate hormonell bedingter Krebsarten (z. B. Brust- und Prostatakrebs) in den asiatischen Ländern wird zum Teil dem hohen Konsum von Soja-Isoflavonen in den meisten traditionellen asiatischen Diäten zugeschrieben.72,73
Isoflavone und Isoflavonoide sind Moleküle, die in vielen Pflanzen, insbesondere in der Bohnenfamilie, vorkommen. Pflanzen verwenden sie als Mittel zur Bekämpfung von Krankheiten und als Stimulanzien für die symbiotischen Bakterien, die ihnen helfen, Stickstoff zum Aufbau von Proteinen zu verwenden. Isoflavone aus Sojabohnen haben auch beim Menschen viele positive und schützende Wirkungen. Die niedrige Rate hormonell bedingter Krebsarten (z. B. Brust- und Prostatakrebs) in den asiatischen Ländern wird zum Teil dem hohen Konsum von Soja-Isoflavonen in den meisten traditionellen asiatischen Diäten zugeschrieben.72,73
Soja-Isoflavone wie Genistein sind starke Modulatoren für NF-kB.73 Genistein wirkt sowohl durch die Aktivierung von NF-kB in den Zellkern, um Entzündungsgene zu aktivieren, als auch durch die Verhinderung früherer molekularer Ereignisse, die zu seiner Aktivierung führen würden.74 Als Folge der Herunterregulierung von NF-kB fördert Genistein den Zelltod von Krebszellen, Apoptose.74
Die Isoflavon-Blockade von NF-kB-Effekten trägt zur Prävention und Modulation von Kolon75 und Lungenkrebs bei.76 Durch die Behandlung mit Isoflavon werden bestimmte Krebszellen zunehmend anfälliger für Bestrahlungs- und Chemotherapieeffekte, vor allem, weil eine verminderte NF-kB-Aktivität die Überlebenssignalwege der Krebszellen beeinträchtigt. 73,77,78
Omega-3-Fettsäuren
 Die an Omega-3-Fettsäuren reichen Diäten sind mit einem verminderten Risiko für verschiedene Arten von Krebs und anderen chronischen Erkrankungen assoziiert.79,80 Die Omega-3-Fettsäuren sind aufgrund ihrer Wirkung auf die wichtigen entzündungsfördernden Enzyme wie entzündungshemmende Mittel COX-2 und 5-LOX bekannt.81Neuere Forschungen zeigen jedoch auch, dass Omega-3-Fettsäuren die Aktivierung von NF-kB durch mehrere Mechanismen hemmen können, wodurch die Entzündung, die zu Krebserkrankungen führen kann, noch stärker unter Kontrolle gebracht werden kann.
Die an Omega-3-Fettsäuren reichen Diäten sind mit einem verminderten Risiko für verschiedene Arten von Krebs und anderen chronischen Erkrankungen assoziiert.79,80 Die Omega-3-Fettsäuren sind aufgrund ihrer Wirkung auf die wichtigen entzündungsfördernden Enzyme wie entzündungshemmende Mittel COX-2 und 5-LOX bekannt.81Neuere Forschungen zeigen jedoch auch, dass Omega-3-Fettsäuren die Aktivierung von NF-kB durch mehrere Mechanismen hemmen können, wodurch die Entzündung, die zu Krebserkrankungen führen kann, noch stärker unter Kontrolle gebracht werden kann.
Omega-3-Fettsäuren sind jetzt mit einer NF-kB-bedingten Verringerung des entzündungsvermittelten Wachstums von Pankreas-, Brust-, Prostata-, Darm- und Hautkrebszellen assoziiert.82-90 Bei Pankreaskarzinomen bewahrte der Omega-3-EPA die Integrität des natürlichen Inhibitors von NF-kB.82Omega-3-Fettsäuren, die in Kombination mit einer konventionellen Chemotherapie eingesetzt werden, wirken synergistisch, um Tumorzellen abzutöten, hauptsächlich durch NF-kB-Hemmung, die die Apoptose fördert.89,91
Zusammenfassung
Eine im letzten Jahr veröffentlichte Durchbruchstudie zeigte, dass eine Entzündung notwendig ist, damit sich Brustkrebszellen vermehren und metastasieren können. Der Master Control-Komplex namens Nuclear Factor-kappaB oder NF-kB löst und reguliert diese Entzündungskaskade, die mit dem Beginn und der Entwicklung von 95% aller Krebserkrankungen in Verbindung steht.
NF-kB stimuliert Gene zur Produktion entzündlicher Zytokine und anderer Signalmoleküle, die das Wachstum und die Entwicklung von Krebs fördern. Die Hemmung von NF-kB ist ein wirksames Ziel für die Krebsprävention und es wurde gezeigt, dass es krebsstimulierende Ereignisse auf allen Ebenen der Karzinogenese (Krebsentwicklung) verringert. Eine breite Palette natürlicher diätetischer Interventionen hemmt die pathologische Aktivität von NF-kB auf kraftvolle Weise. Dies ist eine wichtige Strategie im Kampf gegen Krebs. Zahlreiche neue Studien beschreiben die Auswirkungen der NF-kB-Hemmung auf die Krebsprävention. Die niedrigen Krebsraten in Ländern, in denen natürliche NF-kB-Inhibitoren über die Nahrung aufgenommen werden, weisen nachdrücklich auf den Wert solcher Wirkstoffe als funktionelle Nahrungsmittel und Nutrazeutika hin.
Zusätzlich zu den vielen hier beschriebenen natürlichen Produkten gibt es nun deutliche Hinweise darauf, dass Vitamin D, Withania somnifera (Ashwagandha) und Granatapfelextrakte ähnliche NF-kB-Blockierungsfunktionen aufweisen.102-104
Die gute Nachricht für Life Extension-Mitglieder ist, dass sie seit Jahrzehnten starke NF-kB-hemmende Nährstoffe verwenden.
Mit Genehmigung von Life Extension verwendetes Material. Alle Rechte vorbehalten.
[1] Shin SR, Sanchez-Velar N, Sherr DH, Sonenshein GE. 7,12-dimethylbenz(a)anthracene treatment of a c-rel mouse mammary tumor cell line induces epithelial to mesenchymal transition via activation of nuclear factor-kappaB. Cancer Res. 2006 Mar 1;66(5):2570-5.
Renehan AG, Roberts DL, Dive C. Obesity and cancer: pathophysiological and biological mechanisms. Arch Physiol Biochem. 2008 Feb;114(1):71-83.
Enwonwu CO, Meeks VI. Bionutrition and oral cancer in humans. Crit Rev Oral Biol Med. 1995;6(1):5-17.
[2] Liu M, Sakamaki T, Casimiro MC, et al. The canonical NF-kappaB pathway governs mammary tumorigenesis in transgenic mice and tumor stem cell expansion. Cancer Res. 2010 Dec 15;70(24):10464-73.
[3] Prasad S, Ravindran J, Aggarwal BB. NF-kappaB and cancer: how intimate is this relationship. Mol Cell Biochem. 2010 Mar;336(1-2):25-37.
[4] Pianetti S, Arsura M, Romieu-Mourez R, Coffey RJ, Sonenshein GE. Her-2/neu overexpression induces NF-kappaB via a PI3-kinase/Akt pathway involving calpain-mediated degradation of IkappaB-alpha that can be inhibited by the tumor suppressor PTEN. Oncogene. 2001 Mar 15;20(11):1287-99.
[5] Bhat-Nakshatri P, Sweeney CJ, Nakshatri H. Identification of signal transduction pathways involved in constitutive NF-kappaB activation in breast cancer cells. Oncogene. 2002 Mar 27;21(13):2066-78.
[6] Ahmed KM, Cao N, Li JJ. HER-2 and NF-kappaB as the targets for therapy-resistant breast cancer. Anticancer Res. 2006 Nov-Dec;26(6B):4235-43.
[7] Weldon CB, Burow ME, Rolfe KW, Clayton JL, Jaffe BM, Beckman BS. NF-kappa B-mediated chemoresistance in breast cancer cells. Surgery. 2001 Aug;130(2):143-50.
[8] Plummer SM, Holloway KA, Manson MM, et al. Inhibition of cyclo-oxygenase 2 expression in colon cells by the chemopreventive agent curcumin involves inhibition of NF-kappaB activation via the NIK/IKK signalling complex. Oncogene. 1999 Oct 28;18(44):6013-20.
[9] Dhanalakshmi S, Singh RP, Agarwal C, Agarwal R. Silibinin inhibits constitutive and TNFalpha-induced activation of NF-kappaB and sensitizes human prostate carcinoma DU145 cells to TNFalpha-induced apoptosis. Oncogene. 2002 Mar 7;21(11):1759-67.
[10] Gukovsky I, Reyes CN, Vaquero EC, Gukovskaya AS, Pandol SJ. Curcumin ameliorates ethanol and nonethanol experimental pancreatitis. Am J Physiol Gastrointest Liver Physiol. 2003 Jan;284(1):G85-95.
[11] Pianetti S, Arsura M, Romieu-Mourez R, Coffey RJ, Sonenshein GE. Her-2/neu overexpression induces NF-kappaB via a PI3-kinase/Akt pathway involving calpain-mediated degradation of IkappaB-alpha that can be inhibited by the tumor suppressor PTEN. Oncogene. 2001 Mar 15;20(11):1287-99.
[12] Ahmed KM, Cao N, Li JJ. HER-2 and NF-kappaB as the targets for therapy-resistant breast cancer. Anticancer Res. 2006 Nov-Dec;26(6B):4235-43.
Li SH, Hawthorne VS, Neal CL, et al. Upregulation of neutrophil gelatinase-associated lipocalin by ErbB2 through nuclear factor-kappaB activation. Cancer Res. 2009 Dec 15;69(24):9163-8.
[13] Aravindaram K, Yang NS. Anti-inflammatory plant natural products for cancer therapy. Planta Med. 2010 Aug;76(11):1103-17.
[14] Aravindaram K, Yang NS. Anti-inflammatory plant natural products for cancer therapy. Planta Med. 2010 Aug;76(11):1103-17.
Reuter S, Prasad S, Phromnoi K, et al. Thiocolchicoside exhibits anticancer effects through downregulation of NF-{kappa}B pathway and its regulated gene products linked to inflammation and cancer. Cancer Prev Res (Phila). 2010 Nov;3(11):1462-72.
[15] Ralhan R, Pandey MK, Aggarwal BB. Nuclear factor-kappa B links carcinogenic and chemopreventive agents. Front Biosci (Schol Ed). 2009;1:45-60.
Aggarwal BB, Shishodia S. Suppression of the nuclear factor-kappaB activation pathway by spice-derived phytochemicals: reasoning for seasoning. Ann N Y Acad Sci. 2004 Dec;1030:434-41.
[16] McCarty MF. Targeting multiple signaling pathways as a strategy for managing prostate cancer: multifocal signal modulation therapy. Integr Cancer Ther. 2004 Dec;3(4):349-80.
[17] Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. 2006 Mar 10;160(1):1-40.
[18] Aravindaram K, Yang NS. Anti-inflammatory plant natural products for cancer therapy. Planta Med. 2010 Aug;76(11):1103-17.
[19] Bouzyk E, Gradzka I, Iwanenko T, Kruszewski M, Sochanowicz B, Szumiel I. The response of L5178Y lymphoma sublines to oxidative stress: antioxidant defence, iron content and nuclear translocation of the p65 subunit of NF-kappaB. Acta Biochim Pol. 2000;47(4):881-8.
Saxena A, Saxena AK, Singh J, Bhushan S. Natural antioxidants synergistically enhance the anticancer potential of AP9-cd, a novel lignan composition from Cedrus deodara in human leukemia HL-60 cells. Chem Biol Interact. 2010 Dec 5;188(3):580-90.
[20] Aravindaram K, Yang NS. Anti-inflammatory plant natural products for cancer therapy. Planta Med. 2010 Aug;76(11):1103-17.
Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. 2006 Mar 10;160(1):1-40.
Guo W, Kong E, Meydani M. Dietary polyphenols, inflammation, and cancer. Nutr Cancer. 2009 Nov;61(6):807-10.
Ho HH, Chang CS, Ho WC, Liao SY, Wu CH, Wang CJ. Anti-metastasis effects of gallic acid on gastric cancer cells involves inhibition of NF-kappaB activity and downregulation of PI3K/AKT/small GTPase signals. Food Chem Toxicol. 2010 Aug-Sep;48(8-9):2508-16.
[21] Lin Y, Shi R, Wang X, Shen HM. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr Cancer Drug Targets. 2008 Nov;8(7):634-46.
[22] Prasad AS, Beck FW, Snell DC, Kucuk O. Zinc in cancer prevention. Nutr Cancer. 2009 Nov;61(6):879-87.
[23] Deep G, Gangar SC, Oberlies NH, Kroll DJ, Agarwal R. Isosilybin A induces apoptosis in human prostate cancer cells via targeting Akt, NF-kappaB, and androgen receptor signaling. Mol Carcinog. 2010 Oct;49(10):902-12.
Li L, Zeng J, Gao Y, He D. Targeting silibinin in the antiproliferative pathway. Expert Opin Investig Drugs. 2010 Feb;19(2):243-55.
[24] Palozza P, Colangelo M, Simone R, et al. Lycopene induces cell growth inhibition by altering mevalonate pathway and Ras signaling in cancer cell lines. Carcinogenesis. 2010 Oct;31(10):1813-21.
[25] Kunnumakkara AB, Sung B, Ravindran J, et al. {Gamma}-tocotrienol inhibits pancreatic tumors and sensitizes them to gemcitabine treatment by modulating the inflammatory microenvironment. Cancer Res. 2010 Nov 1;70(21):8695-705.
[26] Palozza P, Colangelo M, Simone R, et al. Lycopene induces cell growth inhibition by altering mevalonate pathway and Ras signaling in cancer cell lines. Carcinogenesis. 2010 Oct;31(10):1813-21.
[27] Duvoix A, Blasius R, Delhalle S, et al. Chemopreventive and therapeutic effects of curcumin. Cancer Lett. 2005 Jun 8;223(2):181-90.
[28] Thangapazham RL, Sharma A, Maheshwari RK. Multiple molecular targets in cancer chemoprevention by curcumin. AAPS J. 2006;8(3):E443-9.
[29] Thangapazham RL, Sharma A, Maheshwari RK. Multiple molecular targets in cancer chemoprevention by curcumin. AAPS J. 2006;8(3):E443-9.
[30] Kunnumakkara AB, Anand P, Aggarwal BB. Curcumin inhibits proliferation, invasion, angiogenesis and metastasis of different cancers through interaction with multiple cell signaling proteins. Cancer Lett. 2008 Oct 8;269(2):199-225.
[31] Reuter S, Eifes S, Dicato M, Aggarwal BB, Diederich M. Modulation of anti-apoptotic and survival pathways by curcumin as a strategy to induce apoptosis in cancer cells. Biochem Pharmacol. 2008 Dec 1;76(11):1340
[32] Everett PC, Meyers JA, Makkinje A, Rabbi M, Lerner A. Preclinical assessment of curcumin as a potential therapy for B-CLL. Am J Hematol. 2007 Jan;82(1):23-30.
Dhillon N, Aggarwal BB, Newman RA, et al. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin Cancer Res. 2008 Jul 15;14(14):4491-9.
Kamat AM, Tharakan ST, Sung B, Aggarwal BB. Curcumin potentiates the antitumor effects of Bacillus Calmette-Guerin against bladder cancer through the downregulation of NF-kappaB and upregulation of TRAIL receptors. Cancer Res. 2009 Dec 1;69(23):8958-66.
Majumdar AP, Banerjee S, Nautiyal J, et al. Curcumin synergizes with resveratrol to inhibit colon cancer. Nutr Cancer. 2009;61(4):544-53.
Bisht S, Mizuma M, Feldmann G, et al. Systemic administration of polymeric nanoparticle-encapsulated curcumin (NanoCurc) blocks tumor growth and metastases in preclinical models of pancreatic cancer. Mol Cancer Ther. 2010 Aug;9(8):2255-64.
Jutooru I, Chadalapaka G, Lei P, Safe S. Inhibition of NFkappaB and pancreatic cancer cell and tumor growth by curcumin is dependent on specificity protein down-regulation. J Biol Chem. 2010 Aug 13;285(33):25332-44.
Ramachandran C, Resek AP, Escalon E, Aviram A, Melnick SJ. Potentiation of gemcitabine by Turmeric Force in pancreatic cancer cell lines. Oncol Rep. 2010 Jun;23(6):1529-35.
Suphim B, Prawan A, Kukongviriyapan U, Kongpetch S, Buranrat B, Kukongviriyapan V. Redox modulation and human bile duct cancer inhibition by curcumin. Food Chem Toxicol. 2010 Aug-Sep;48(8-9):2265-72.
Zhang C, Li B, Zhang X, Hazarika P, Aggarwal BB, Duvic M. Curcumin selectively induces apoptosis in cutaneous T-cell lymphoma cell lines and patients’ PBMCs: potential role for STAT-3 and NF-kappaB signaling. J Invest Dermatol. 2010 Aug;130(8):2110-9.
[33] Aggarwal BB. Targeting inflammation-induced obesity and metabolic diseases by curcumin and other nutraceuticals. Annu Rev Nutr. 2010 Aug 21;30:173-99.
[34] Dong G, Chen Z, Kato T, Van Waes C. The host environment promotes the constitutive activation of nuclear factor-kappaB and proinflammatory cytokine expression during metastatic tumor progression of murine squamous cell carcinoma. Cancer Res. 1999 Jul 15;59(14):3495-504.
[35] Pianetti S, Arsura M, Romieu-Mourez R, Coffey RJ, Sonenshein GE. Her-2/neu overexpression induces NF-kappaB via a PI3-kinase/Akt pathway involving calpain-mediated degradation of IkappaB-alpha that can be inhibited by the tumor suppressor PTEN. Oncogene. 2001 Mar 15;20(11):1287-99.
Guo W, Kong E, Meydani M. Dietary polyphenols, inflammation, and cancer. Nutr Cancer. 2009 Nov;61(6):807-10.
[36] Pikarsky E, Porat RM, Stein I, et al. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature. 2004 Sep 23;431(7007):461-6.
[37] Bhat-Nakshatri P, Sweeney CJ, Nakshatri H. Identification of signal transduction pathways involved in constitutive NF-kappaB activation in breast cancer cells. Oncogene. 2002 Mar 27;21(13):2066-78.
[38] Bhat-Nakshatri P, Sweeney CJ, Nakshatri H. Identification of signal transduction pathways involved in constitutive NF-kappaB activation in breast cancer cells. Oncogene. 2002 Mar 27;21(13):2066-78.
[39] Bhat-Nakshatri P, Sweeney CJ, Nakshatri H. Identification of signal transduction pathways involved in constitutive NF-kappaB activation in breast cancer cells. Oncogene. 2002 Mar 27;21(13):2066-78.
[40] Chapkin RS, Kim W, Lupton JR, McMurray DN. Dietary docosahexaenoic and eicosapentaenoic acid: emerging mediators of inflammation. Prostaglandins Leukot Essent Fatty Acids. 2009 Aug-Sep;81(2-3):187-91.
[41] Borek C. Antioxidant health effects of aged garlic extract. J Nutr. 2001 Mar;131(3s):1010S-5S.
[42] Ban JO, Lee HS, Jeong HS, et al. Thiacremonone augments chemotherapeutic agent-induced growth inhibition in human colon cancer cells through inactivation of nuclear factor-{kappa}B. Mol Cancer Res. 2009 Jun;7(6):870-9.
Pratheeshkumar P, Thejass P, Kutan G. Diallyl disulfide induces caspase-dependent apoptosis via mitochondria-mediated intrinsic pathway in B16F-10 melanoma cells by up-regulating p53, caspase-3 and down-regulating pro-inflammatory cytokines and nuclear factor-kappabeta-mediated Bcl-2 activation. J Environ Pathol Toxicol Oncol. 2010;29(2):113-25.
[43] Ban JO, Lee HS, Jeong HS, et al. Thiacremonone augments chemotherapeutic agent-induced growth inhibition in human colon cancer cells through inactivation of nuclear factor-{kappa}B. Mol Cancer Res. 2009 Jun;7(6):870-9.
Pratheeshkumar P, Thejass P, Kutan G. Diallyl disulfide induces caspase-dependent apoptosis via mitochondria-mediated intrinsic pathway in B16F-10 melanoma cells by up-regulating p53, caspase-3 and down-regulating pro-inflammatory cytokines and nuclear factor-kappabeta-mediated Bcl-2 activation. J Environ Pathol Toxicol Oncol. 2010;29(2):113-25.
Ban JO, Yuk DY, Woo KS, et al. Inhibition of cell growth and induction of apoptosis via inactivation of NF-kappaB by a sulfurcompound isolated from garlic in human colon cancer cells. J Pharmacol Sci. 2007 Aug;104(4):374-83.
Karmakar S, Banik NL, Patel SJ, Ray SK. Garlic compounds induced calpain and intrinsic caspase cascade for apoptosis in human malignant neuroblastoma SH-SY5Y cells. Apoptosis. 2007 Apr;12(4):671-84.
Sriram N, Kalayarasan S, Ashokkumar P, Sureshkumar A, Sudhandiran G. Diallyl sulfide induces apoptosis in Colo 320 DM human colon cancer cells: involvement of caspase-3, NF-kappaB, and ERK-2. Mol Cell Biochem. 2008 Apr;311(1-2):157-65.
[44] Ban JO, Yuk DY, Woo KS, et al. Inhibition of cell growth and induction of apoptosis via inactivation of NF-kappaB by a sulfurcompound isolated from garlic in human colon cancer cells. J Pharmacol Sci. 2007 Aug;104(4):374-83.
Sriram N, Kalayarasan S, Ashokkumar P, Sureshkumar A, Sudhandiran G. Diallyl sulfide induces apoptosis in Colo 320 DM human colon cancer cells: involvement of caspase-3, NF-kappaB, and ERK-2. Mol Cell Biochem. 2008 Apr;311(1-2):157-65.
[45] Kim SO, Kundu JK, Shin YK, et al. [6]-Gingerol inhibits COX-2 expression by blocking the activation of p38 MAP kinase and NF-kappaB in phorbol ester-stimulated mouse skin. Oncogene. 2005 Apr 7;24(15):2558-67.
[46] Oyagbemi AA, Saba AB, Azeez OI. Molecular targets of [6]-gingerol: Its potential roles in cancer chemoprevention. Biofactors. 2010 May-Jun;36(3):169-78.
[47] Kim SO, Kundu JK, Shin YK, et al. [6]-Gingerol inhibits COX-2 expression by blocking the activation of p38 MAP kinase and NF-kappaB in phorbol ester-stimulated mouse skin. Oncogene. 2005 Apr 7;24(15):2558-67.
Oyagbemi AA, Saba AB, Azeez OI. Molecular targets of [6]-gingerol: Its potential roles in cancer chemoprevention. Biofactors. 2010 May-Jun;36(3):169-78.
[48] Takada Y, Murakami A, Aggarwal BB. Zerumbone abolishes NF-kappaB and IkappaBalpha kinase activation leading to suppression of antiapoptotic and metastatic gene expression, upregulation of apoptosis, and downregulation of invasion. Oncogene. 2005 Oct 20;24(46):6957-69.
[49] Ishiguro K, Ando T, Maeda O, et al. Ginger ingredients reduce viability of gastric cancer cells via distinct mechanisms. Biochem Biophys Res Commun. 2007 Oct 12;362(1):218-23.
Kim JK, Kim Y, Na KM, Surh YJ, Kim TY. [6]-Gingerol prevents UVB-induced ROS production and COX-2 expression in vitro and in vivo. Free Radic Res. 2007 May;41(5):603-14.
Rhode J, Fogoros S, Zick S, et al. Ginger inhibits cell growth and modulates angiogenic factors in ovarian cancer cells. BMC Complement Altern Med. 2007;7:44.
Habib SH, Makpol S, Abdul Hamid NA, Das S, Ngah WZ, Yusof YA. Ginger extract (Zingiber officinale) has anti-cancer and anti-inflammatory effects on ethionine-induced hepatoma rats. Clinics (Sao Paulo). 2008 Dec;63(6):807-13.
Kim M, Miyamoto S, Yasui Y, Oyama T, Murakami A, Tanaka T. Zerumbone, a tropical ginger sesquiterpene, inhibits colon and lung carcinogenesis in mice. Int J Cancer. 2009 Jan 15;124(2):264-71.
Sung B, Murakami A, Oyajobi BO, Aggarwal BB. Zerumbone abolishes RANKL-induced NF-kappaB activation, inhibits osteoclastogenesis, and suppresses human breast cancer-induced bone loss in athymic nude mice. Cancer Res. 2009 Feb 15;69(4):1477-84.
[50] Prasad S, Ravindran J, Aggarwal BB. NF-kappaB and cancer: how intimate is this relationship. Mol Cell Biochem. 2010 Mar;336(1-2):25-37.
[51] Shin SR, Sanchez-Velar N, Sherr DH, Sonenshein GE. 7,12-dimethylbenz(a)anthracene treatment of a c-rel mouse mammary tumor cell line induces epithelial to mesenchymal transition via activation of nuclear factor-kappaB. Cancer Res. 2006 Mar 1;66(5):2570-5.
Renehan AG, Roberts DL, Dive C. Obesity and cancer: pathophysiological and biological mechanisms. Arch Physiol Biochem. 2008 Feb;114(1):71-83.
Enwonwu CO, Meeks VI. Bionutrition and oral cancer in humans. Crit Rev Oral Biol Med. 1995;6(1):5-17.
[52] Shin SR, Sanchez-Velar N, Sherr DH, Sonenshein GE. 7,12-dimethylbenz(a)anthracene treatment of a c-rel mouse mammary tumor cell line induces epithelial to mesenchymal transition via activation of nuclear factor-kappaB. Cancer Res. 2006 Mar 1;66(5):2570-5.
Pratt MA, Tibbo E, Robertson SJ, et al. The canonical NF-kappaB pathway is required for formation of luminal mammary neoplasias and is activated in the mammary progenitor population. Oncogene. 2009 Jul 30;28(30):2710-22.
[53] Pianetti S, Arsura M, Romieu-Mourez R, Coffey RJ, Sonenshein GE. Her-2/neu overexpression induces NF-kappaB via a PI3-kinase/Akt pathway involving calpain-mediated degradation of IkappaB-alpha that can be inhibited by the tumor suppressor PTEN. Oncogene. 2001 Mar 15;20(11):1287-99.
Li SH, Hawthorne VS, Neal CL, et al. Upregulation of neutrophil gelatinase-associated lipocalin by ErbB2 through nuclear factor-kappaB activation. Cancer Res. 2009 Dec 15;69(24):9163-8.
[54] deGraffenried LA, Chandrasekar B, Friedrichs WE, et al. NF-kappa B inhibition markedly enhances sensitivity of resistant breast cancer tumor cells to tamoxifen. Ann Oncol. 2004 Jun;15(6):885-90.
[55] Brantley DM, Chen CL, Muraoka RS, et al. Nuclear factor-kappaB (NF-kappaB) regulates proliferation and branching in mouse mammary epithelium. Mol Biol Cell. 2001 May;12(5):1445-55.
[56] deGraffenried LA, Chandrasekar B, Friedrichs WE, et al. NF-kappa B inhibition markedly enhances sensitivity of resistant breast cancer tumor cells to tamoxifen. Ann Oncol. 2004 Jun;15(6):885-90.
Hagemann T, Wilson J, Kulbe H, et al. Macrophages induce invasiveness of epithelial cancer cells via NF-kappa B and JNK. J Immunol. 2005 Jul 15;175(2):1197-205.
Liu M, Ju X, Willmarth NE, et al. Nuclear factor-kappaB enhances ErbB2-induced mammary tumorigenesis and neoangiogenesis in vivo. Am J Pathol. 2009 May;174(5):1910-20.
Beshir AB, Ren G, Magpusao AN, Barone LM, Yeung KC, Fenteany G. Raf kinase inhibitor protein suppresses nuclear factor-kappaB-dependent cancer cell invasion through negative regulation of matrix metalloproteinase expression. Cancer Lett. 2010 Dec 28;299(2):137-49.
[57] Tapia MA, Gonzalez-Navarrete I, Dalmases A, et al. Inhibition of the canonical IKK/NF kappa B pathway sensitizes human cancer cells to doxorubicin. Cell Cycle. 2007 Sep 15;6(18):2284-92.
[58] Prasad S, Ravindran J, Aggarwal BB. NF-kappaB and cancer: how intimate is this relationship. Mol Cell Biochem. 2010 Mar;336(1-2):25-37.
[59] Ahmed KM, Cao N, Li JJ. HER-2 and NF-kappaB as the targets for therapy-resistant breast cancer. Anticancer Res. 2006 Nov-Dec;26(6B):4235-43.
Li SH, Hawthorne VS, Neal CL, et al. Upregulation of neutrophil gelatinase-associated lipocalin by ErbB2 through nuclear factor-kappaB activation. Cancer Res. 2009 Dec 15;69(24):9163-8.
[60] Pikarsky E, Porat RM, Stein I, et al. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature. 2004 Sep 23;431(7007):461-6.
[61] Yang CS, Maliakal P, Meng X. Inhibition of carcinogenesis by tea. Annu Rev Pharmacol Toxicol. 2002;42:25-54.
Butt MS, Sultan MT. Green tea: nature’s defense against malignancies. Crit Rev Food Sci Nutr. 2009 May;49(5):463-73.
[62] Khan N, Afaq F, Saleem M, Ahmad N, Mukhtar H. Targeting multiple signaling pathways by green tea polyphenol (-)-epigallocatechin-3-gallate. Cancer Res. 2006 Mar 1;66(5):2500-5.
Khan N, Mukhtar H. Multitargeted therapy of cancer by green tea polyphenols. Cancer Lett. 2008 Oct 8;269(2):269-80.
[63] Khan N, Afaq F, Saleem M, Ahmad N, Mukhtar H. Targeting multiple signaling pathways by green tea polyphenol (-)-epigallocatechin-3-gallate. Cancer Res. 2006 Mar 1;66(5):2500-5.
Khan N, Mukhtar H. Multitargeted therapy of cancer by green tea polyphenols. Cancer Lett. 2008 Oct 8;269(2):269-80.
Navarro-Peran E, Cabezas-Herrera J, Sanchez-Del-Campo L, Garcia-Canovas F, Rodriguez-Lopez JN. The anti-inflammatory and anti-cancer properties of epigallocatechin-3-gallate are mediated by folate cycle disruption, adenosine release and NF-kappaB suppression. Inflamm Res. 2008 Oct;57(10):472-8.
[64] Horia E, Watkins BA. Complementary actions of docosahexaenoic acid and genistein on COX-2, PGE2 and invasiveness in MDA-MB-231 breast cancer cells. Carcinogenesis. 2007 Apr;28(4):809-15.
[65] Siddiqui IA, Shukla Y, Adhami VM, et al. Suppression of NFkappaB and its regulated gene products by oral administration of green tea polyphenols in an autochthonous mouse prostate cancer model. Pharm Res. 2008 Sep;25(9):2135-42.
Amin AR, Khuri FR, Chen ZG, Shin DM. Synergistic growth inhibition of squamous cell carcinoma of the head and neck by erlotinib and epigallocatechin-3-gallate: the role of p53-dependent inhibition of nuclear factor-kappaB. Cancer Prev Res (Phila). 2009 Jun;2(6):538-45.
Nihal M, Ahsan H, Siddiqui IA, Mukhtar H, Ahmad N, Wood GS. (-)-Epigallocatechin-3-gallate (EGCG) sensitizes melanoma cells to interferon induced growth inhibition in a mouse model of human melanoma. Cell Cycle. 2009 Jul 1;8(13):2057-63.
Sen T, Moulik S, Dutta A, et al. Multifunctional effect of epigallocatechin-3-gallate (EGCG) in downregulation of gelatinase-A (MMP-2) in human breast cancer cell line MCF-7. Life Sci. 2009 Feb 13;84(7-8):194-204.
Roy P, Nigam N, Singh M, et al. Tea polyphenols inhibit cyclooxygenase-2 expression and block activation of nuclear factor-kappa B and Akt in diethylnitrosoamine induced lung tumors in Swiss mice. Invest New Drugs. 2010 Aug;28(4):466-71.
[66] Sarkar FH, Li Y. The role of isoflavones in cancer chemoprevention. Front Biosci. 2004 Sep 1;9:2714-24.
Banerjee S, Li Y, Wang Z, Sarkar FH. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008 Oct 8;269(2):226-42.
[67] Banerjee S, Li Y, Wang Z, Sarkar FH. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008 Oct 8;269(2):226-42.
[68] Li Z, Li J, Mo B, et al. Genistein induces cell apoptosis in MDA-MB-231 breast cancer cells via the mitogen-activated protein kinase pathway. Toxicol In Vitro. 2008 Oct;22(7):1749-53.
[69] Li Z, Li J, Mo B, et al. Genistein induces cell apoptosis in MDA-MB-231 breast cancer cells via the mitogen-activated protein kinase pathway. Toxicol In Vitro. 2008 Oct;22(7):1749-53.
[70] Ravasco P, Aranha MM, Borralho PM, et al. Colorectal cancer: can nutrients modulate NF-kappaB and apoptosis? Clin Nutr. 2010 Feb;29(1):42-6.
Gadgeel SM, Ali S, Philip PA, Wozniak A, Sarkar FH. Genistein enhances the effect of epidermal growth factor receptor tyrosine kinase inhibitors and inhibits nuclear factor kappa B in nonsmall cell lung cancer cell lines. Cancer. 2009 May 15;115(10):2165-76.
[71] Banerjee S, Li Y, Wang Z, Sarkar FH. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008 Oct 8;269(2):226-42.
Singh-Gupta V, Zhang H, Banerjee S, et al. Radiation-induced HIF-1alpha cell survival pathway is inhibited by soy isoflavones in prostate cancer cells. Int J Cancer. 2009 Apr 1;124(7):1675-84.
Zhang B, Shi ZL, Liu B, Yan XB, Feng J, Tao HM. Enhanced anticancer effect of gemcitabine by genistein in osteosarcoma: the role of Akt and nuclear factor-kappaB. Anticancer Drugs. 2010 Mar;21(3):288-96.
[72] Slagsvold JE, Pettersen CH, Storvold GL, Follestad T, Krokan HE, Schonberg SA. DHA alters expression of target proteins of cancer therapy in chemotherapy resistant SW620 colon cancer cells. Nutr Cancer. 2010 Jul;62(5):611-21.
Jho DH, Cole SM, Lee EM, Espat NJ. Role of omega-3 fatty acid supplementation in inflammation and malignancy. Integr Cancer Ther. 2004 Jun;3(2):98-111.
[73] Groeger AL, Cipollina C, Cole MP, et al. Cyclooxygenase-2 generates anti-inflammatory mediators from omega-3 fatty acids. Nat Chem Biol. 2010 Jun;6(6):433-41.
[74] Ross JA, Maingay JP, Fearon KC, Sangster K, Powell JJ. Eicosapentaenoic acid perturbs signalling via the NFkappaB transcriptional pathway in pancreatic tumour cells. Int J Oncol. 2003 Dec;23(6):1733-8.
Narayanan NK, Narayanan BA, Reddy BS. A combination of docosahexaenoic acid and celecoxib prevents prostate cancer cell growth in vitro and is associated with modulation of nuclear factor-kappaB, and steroid hormone receptors. Int J Oncol. 2005 Mar;26(3):785-92.
Schley PD, Jijon HB, Robinson LE, Field CJ. Mechanisms of omega-3 fatty acid-induced growth inhibition in MDA-MB-231 human breast cancer cells. Breast Cancer Res Treat. 2005 Jul;92(2):187-95.
Hering J, Garrean S, Dekoj TR, et al. Inhibition of proliferation by omega-3 fatty acids in chemoresistant pancreatic cancer cells. Ann Surg Oncol. 2007 Dec;14(12):3620-8.
Horia E, Watkins BA. Complementary actions of docosahexaenoic acid and genistein on COX-2, PGE2 and invasiveness in MDA-MB-231 breast cancer cells. Carcinogenesis. 2007 Apr;28(4):809-15.
Nowak J, Weylandt KH, Habbel P, et al. Colitis-associated colon tumorigenesis is suppressed in transgenic mice rich in endogenous n-3 fatty acids. Carcinogenesis. 2007 Sep;28(9):1991-5.
Kim HH, Lee Y, Eun HC, Chung JH. Eicosapentaenoic acid inhibits TNF-alpha-induced matrix metalloproteinase-9 expression in human keratinocytes, HaCaT cells. Biochem Biophys Res Commun. 2008 Apr 4;368(2):343-9.
Shaikh IA, Brown I, Schofield AC, Wahle KW, Heys SD. Docosahexaenoic acid enhances the efficacy of docetaxel in prostate cancer cells by modulation of apoptosis: the role of genes associated with the NF-kappaB pathway. Prostate. 2008 Nov 1;68(15):1635-46.
Ghosh-Choudhury T, Mandal CC, Woodruff K, et al. Fish oil targets PTEN to regulate NFkappaB for downregulation of anti-apoptotic genes in breast tumor growth. Breast Cancer Res Treat. 2009 Nov;118(1):213-28.
[75] Ross JA, Maingay JP, Fearon KC, Sangster K, Powell JJ. Eicosapentaenoic acid perturbs signalling via the NFkappaB transcriptional pathway in pancreatic tumour cells. Int J Oncol. 2003 Dec;23(6):1733-8.
[76] Shaikh IA, Brown I, Schofield AC, Wahle KW, Heys SD. Docosahexaenoic acid enhances the efficacy of docetaxel in prostate cancer cells by modulation of apoptosis: the role of genes associated with the NF-kappaB pathway. Prostate. 2008 Nov 1;68(15):1635-46.
Ding WQ, Liu B, Vaught JL, Palmiter RD, Lind SE. Clioquinol and docosahexaenoic acid act synergistically to kill tumor cells. Mol Cancer Ther. 2006 Jul;5(7):1864-72.
[77] Mulabagal V, Subbaraju GV, Rao CV, et al. Withanolide sulfoxide from Aswagandha roots inhibits nuclear transcription factor-kappa-B, cyclooxygenase and tumor cell proliferation. Phytother Res. 2009 Jul;23(7):987-92.
Kwon HJ, Won YS, Suh HW, et al. Vitamin D3 upregulated protein 1 suppresses TNF-alpha-induced NF-kappaB activation in hepatocarcinogenesis. J Immunol. 2010 Oct 1;185(7):3980-9.
Rettig MB, Heber D, An J, et al. Pomegranate extract inhibits androgen-independent prostate cancer growth through a nuclear factor-kappaB-dependent mechanism. Mol Cancer Ther. 2008 Sep;7(9):2662-71.
Zugehörige Erzeugnisse
-

Super Absorbierbare Tocotrienole
Preis:30,25 €Unterstützen gesundes Haarwachstum und mehr Werden aus rotem Palmöl... -

Granatapfel Komplett
Preis:29,28 €Standardisierte Extrakte aus dem gesamten Frucht, Blüten und Samenöl Die... -

Mega Grüntee-Extrakt (entkoffeiniert)
Preis:31,23 €Eine Kapsel enthält mehr Polyphenole als sieben Tassen grünen Tee... -

Zink-Kapseln
Preis:14,13 €Unterstützt die natürliche Immunabwehr Stärkt die natürliche Immunabwehr... -

Curcumin Elite™ Kurkuma-Extrakt
Preis:35,14 €Diese firmeneigene Curcumin-Formel ist die beste, die wir jemals angeboten... -

Super Omega-3 EPA/DHA mit Sesamlignanen und...
Preis:43,64 €Nehmen Sie ausreichend Omega-3-Fettsäuren zu sich Leistungsstarke Hilfe... -

Fortschrittlicher Curcumin Elite ™...
Preis:26,10 €Hoch resorbierbares Curcumin mit ergänzenden Pflanzenextrakten Bietet 45x... -

Optimierter Ashwagandha-Extrakt
Preis:12,83 €Erhöht die mentale Energie und Konzentration Es wird angenommen, dass es...
Ähnliche Artikel
-
 Alterndes Gelenken neu aufbauen!
In diesem Artikel erfahren Sie etwas über UC-II® oder nicht denaturiertes Kollagen Typ II, einen neuartigen Eingriff...
Alterndes Gelenken neu aufbauen!
In diesem Artikel erfahren Sie etwas über UC-II® oder nicht denaturiertes Kollagen Typ II, einen neuartigen Eingriff... -
Ist konventioneller Granatapfel-Extrakt genug?
Zahlreiche klinische Daten haben Granatapfel als eine multimodale Abwehr gegen die häufigsten Todesursachen in der...
-
Nährstoffe der nächsten Generation für die schnelle Linderung von Gelenkschmerzen
In den letzten Jahren mussten sich Erwachsene, die nach den lähmenden Schmerzen der Arthritis lindern wollten,...
-
Liponsäure kehrt den mitochondrialen Zerfall um
Es wird geschätzt, dass 85% des in jedem Atemzug enthaltenen Sauerstoffs von den Mitochondrien in jeder Körperzelle...
-
Vitamin C und Dihydroquercetin
Jeden Tag werden unsere Körper ständig von schädlichen Agenten, die als freie Radikale bekannt sind, angegriffen. Im...