The Inflammatory Factor Underlying Most Cancers
 Implicated in the onset of 95% of all cancers is a common but under-recognized molecule most people have never heard of: nuclear factor-kappaB or NF-kB.1-3 Emerging research is validating the danger it poses to maturing individuals.
Implicated in the onset of 95% of all cancers is a common but under-recognized molecule most people have never heard of: nuclear factor-kappaB or NF-kB.1-3 Emerging research is validating the danger it poses to maturing individuals.
For example, a landmark study4 published late last year confirmed, for the first time, NF-kB’s central role in the onset of malignant breast cancer.
A team of researchers found that breast cancer cells require a specific inflammatory cascade in order to proliferate and metastasize.
As one of the proteins that activates genes involved in inflammation, NF-kB is the master regulator of this pathologic cancer-causing process.
Fortunately, there exists a broad array of nutrients many Life Extension® members already take that have been shown to neutralize and even reverse the cancer-causing inflammatory cascade mediated by NF-kB.
In this article, a comprehensive survey of NF-kB-inhibiting compounds is detailed. You will learn how NF-kB triggers the growth and development of many kinds of tumors, operating as the master switch for cells “pre-loaded” with cancer-causing genes. You’ll also discover compelling evidence about low-cost interventions that suppress NF-kB, switch off lethal cancer genes, and disrupt the inflammatory cascade cancer cells need in order to spread.
Since its discovery in 1986, NF-kB has been known to be closely associated with development of cancer.5 Today we know that most, if not all cancers have abnormally high levels of active NF-kB, which keeps them in a state of sustained inflammation.6 Conversely, certain so-called oncogenes (abnormal genes that can initiate cancer development) act in part by activating NF-kB.7 Together these facts conspire to produce a steady stream of inflammatory and cancer-promoting cytokines within a growing tumor.8
The production of these cell signaling molecules makes it much more likely that a given cancer-prone cell will go on to form a malignant tumor. It can also mean that a given tumor is much more resistant to radiation and chemotherapy, since NF-kB generally promotes cancer cell survival.9
The NF-kB-Cancer Blockade
 NF-kB triggers the cascade of events that link inflammation with cancer development. Once that process has been initiated, NF-kB continues to fan the flames, contributing to cancer cells’ invasiveness, ability to stimulate a fresh blood supply, and ability to spread by metastasis throughout the body.
NF-kB triggers the cascade of events that link inflammation with cancer development. Once that process has been initiated, NF-kB continues to fan the flames, contributing to cancer cells’ invasiveness, ability to stimulate a fresh blood supply, and ability to spread by metastasis throughout the body.
But our knowledge about the damage done by NF-kB can be turned to good. Almost as soon as it was discovered, forward-thinking researchers recognized that if they could block activation of NF-kB, they might break an essential link in cancer development.
We’ve known for nearly 12 years, for example, that we could reduce inflammation in colon cancer cells by treatment with the spice curcumin, a well-known inhibiter of NF-kB activation.10 Blocking NF-kB activation can also sensitize human prostate cancer cells to apoptosis—the programmed cell death that keeps cancers under control.11 And natural molecules that inhibit NF-kB can limit inflammatory changes in the pancreas caused by alcohol.12
The potential for blocking NF-kB offers hope in the context of the deadly HER-2 breast cancer gene. HER-2 cancers are normally highly treatment-resistant and carry a poor prognosis.6 Blocking NF-kB can make the breast cancer cells more amenable to chemotherapy.8,13
Not surprisingly, Big Pharma is intensely interested in discovering new (and profitable) drugs that block NF-kB.14 But nature has provided us with a substantial number of substances that block or inhibit the pro-inflammatory, pro-cancer effects of renegade NF-kB—and they are readily available, affordable, and safe. Let’s look at a sampling.
Nutrients to Quell NF-kB’s Inflammatory, Pro-Cancer Effects
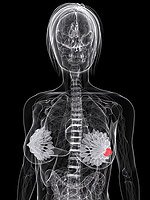 By some estimates, anywhere from 67% to 90% of human cancers could be prevented by modifications to lifestyle, specifically to diet.14,15 Suppression of NF-kB by phytochemicals present in fruits, vegetables, and many spices has in fact been said to provide the molecular basis for their ability to prevent cancer.16,17
By some estimates, anywhere from 67% to 90% of human cancers could be prevented by modifications to lifestyle, specifically to diet.14,15 Suppression of NF-kB by phytochemicals present in fruits, vegetables, and many spices has in fact been said to provide the molecular basis for their ability to prevent cancer.16,17
The following nutraceutical agents can bring us closer to successful chemoprevention through their ability to oppose NF-kB’s pro-inflammatory, pro-cancer effects. Unlike single-targeted drugs, most nutraceuticals are multitargeted. That gives them astonishing efficiency and can augment their effectiveness, especially when acting to interrupt complex signaling pathways like those involved with NF-kB.18
Antioxidants
 Oxidative damage to DNA, proteins, and other vital structures is a major factor in the initiation, promotion, and spread of cancers. That makes antioxidants important agents in the fight to prevent cancer.19 But many antioxidants can also block NF-kB, adding a valuable degree of specificity to their effects. They do this via a number of closely related but distinctive mechanisms.14
Oxidative damage to DNA, proteins, and other vital structures is a major factor in the initiation, promotion, and spread of cancers. That makes antioxidants important agents in the fight to prevent cancer.19 But many antioxidants can also block NF-kB, adding a valuable degree of specificity to their effects. They do this via a number of closely related but distinctive mechanisms.14
One important way of blocking NF-kB activity is to prevent it from becoming activated by moving into the cell nucleus. That denies NF-kB access to the pro-inflammatory genes that it regulates, thereby preventing the inflammatory response before it starts. Cells with greater levels of internal antioxidant defenses generally have less NF-kB activation. Increasing those defenses is correlated with reduced NF-kB expression in cell nuclei.20,21
A large number of antioxidants are under review for specific NF-kB-blocking activity. These include vitamins C and E, carotenoids, the so-called thiol antioxidants (glutathione, thioredoxin, and lipoic acid), flavonoids and polyphenols, selenium, zinc, and others.14,19,22,23 The flavonoid luteolin, for example, can sensitize cancer cells to chemotherapy agents by suppressing a variety of cell-survival pathways, including NF-kB.24 And zinc, an essential trace element, inhibits NF-kB indirectly by inducing a separate regulatory protein called A-20.25
 Milk thistle extracts enhance prostate cancer cell death through apoptosis by reducing levels of NF-kB in the cell nucleus.26,27 The tomato carotenoid pigment lycopene prevents NF-kB activation in prostate cancer by a different mechanism.28 A form of vitamin E called gamma-tocotrienol downregulates NF-kB in pancreatic tumor cells.29 Even certain coffee components, the antioxidant molecules called diterpenes, suppress NF-kB activation.30 And a host of antioxidant compounds derived from black pepper reduce NF-kB activation.31 Finally, the tomato polyphenol lycopene reduces NF-kB activation in a variety of cancer cell types.28
Milk thistle extracts enhance prostate cancer cell death through apoptosis by reducing levels of NF-kB in the cell nucleus.26,27 The tomato carotenoid pigment lycopene prevents NF-kB activation in prostate cancer by a different mechanism.28 A form of vitamin E called gamma-tocotrienol downregulates NF-kB in pancreatic tumor cells.29 Even certain coffee components, the antioxidant molecules called diterpenes, suppress NF-kB activation.30 And a host of antioxidant compounds derived from black pepper reduce NF-kB activation.31 Finally, the tomato polyphenol lycopene reduces NF-kB activation in a variety of cancer cell types.28
Clearly, the list of antioxidant nutrients with NF-kB-suppressing effects is growing rapidly. A good general rule for the present is to make certain that you are including an adequate supply of potent, high-quality antioxidants as part of your health regimen. Let’s now turn to some specific nutrients that have additional NF-kB-blocking effects that can optimize your cancer protection.
- A breakthrough study revealed that inflammation is necessary in order for breast cancer cells to proliferate and metastasize.
- The master control complex called nuclear factor-kappaB or NF-kB triggers and regulates this inflammatory cascade, which has been implicated in the onset and development of 95% of all cancers.
- NF-kB stimulates genes to produce inflammatory cytokines and other signaling molecules that promote cancer growth and development.
- Inhibition of NF-kB is a powerful target for cancer prevention and has been shown to reduce cancer-stimulative events at every level of carcinogenesis (cancer development).
- A broad array of natural dietary interventions powerfully inhibits pathologic NF-kB activity, comprising a critical strategy in the fight against cancer.
Curcumin
 Curcumin is an extraordinary molecule extracted from the spice we know as turmeric. It is among the most potent of the natural inhibitors of NF-kB, and as such has been intensely studied for its anticancer potential.32
Curcumin is an extraordinary molecule extracted from the spice we know as turmeric. It is among the most potent of the natural inhibitors of NF-kB, and as such has been intensely studied for its anticancer potential.32
Curcumin is beneficial at all three major stages of carcinogenesis: initiation, progression, and promotion.33 Most of that benefit has been linked to its inhibition of NF-kB and subsequent inhibition of pro-inflammatory signaling.33 NF-kB inhibition is also credited with curcumin’s ability to inhibit cancer cell proliferation, invasion, new blood vessel formation, and metastasis.34 Another major consequence of NF-kB inhibition by curcumin is to allow the natural course of apoptosis to take place in cancer cells, triggering them to die off rather than multiply and spread.35
Curcumin is showing promise in treating or preventing some of mankind’s most common, and most dangerous, malignancies. These include chronic lymphocytic leukemias, lymphomas, pancreatic cancers, bladder cancer, colon cancer, and cancer of the bile ducts.36-44 And curcumin can help to suppress the cancer-provoking inflammation generated by obesity, one of today’s most pressing health problems.45
 NF-kB is scientifically known as a “transcription factor.”92 It regulates how and when the information in certain human genes becomes translated into new protein molecules.
NF-kB is scientifically known as a “transcription factor.”92 It regulates how and when the information in certain human genes becomes translated into new protein molecules.
Key among those genes are those that promote inflammatory responses, many of which also promote cell growth, survival, and transformation into cancer.6,22 In fact, in the early 2000s it became clear that NF-kB was the “missing link” between inflammation and cancer.93 Of course, that’s not its only role.
In healthy people, NF-kB serves a vital function just by remaining inactive. That helps to keep the powerful cellular machinery of inflammation running just at idle. When a threat arises, however, NF-kB becomes activated and goes to work within the nuclei of cells (hence the term “nuclear factor”).7
A cell’s nucleus is where all the genetic information is stored in the form of DNA in our chromosomes. Under normal circumstances, NF-kB “lives” not in the nucleus, but in the cell’s cytoplasm, the main body of the cell.7 When an infection or other foreign invader threatens, NF-kB moves from the cell’s cytoplasm into its nucleus.7 Once inside the nucleus, NF-kB binds to specific portions of the chromosome, where it activates transcription of a gene into its protein product.
Many genes that come under the control of NF-kB are pro-inflammatory genes, that is, their products are inflammatory cytokines such as interleukins, tumor necrosis factor, and others. Once the inflammatory stimulus goes away, NF-kB normally disengages from the chromosome and returns to the cytoplasm, and the inflammatory gene stops producing its cytokines. In other words, under normal circumstances, inflammation dies away when it is no longer beneficial.94
But that’s only under normal circumstances, and cancer is anything but normal.
Garlic Extracts
 Garlic, like turmeric, has a lengthy history of use in traditional medical systems. When garlic extracts are allowed to age, they stabilize and retain strong antioxidant powers that indirectly down-regulate NF-kB.46 Individual components from such garlic extracts include diallyl sulfide, diallyl disulfide, and thiacremonone, all of which demonstrate direct NF-kB-inhibitory activities.17,47,48
Garlic, like turmeric, has a lengthy history of use in traditional medical systems. When garlic extracts are allowed to age, they stabilize and retain strong antioxidant powers that indirectly down-regulate NF-kB.46 Individual components from such garlic extracts include diallyl sulfide, diallyl disulfide, and thiacremonone, all of which demonstrate direct NF-kB-inhibitory activities.17,47,48
NF-kB inhibition by garlic extracts is showing promise in preventing growth and spread of many tumor types, including colon cancer, malignant neuroblastoma, and melanoma.47-51 Garlic extracts also powerfully induce cancer cell death by apoptosis.49,51
Ginger
 Ginger is a spice from southern Asia, that has many health-giving specific components.52 Gingerol is the chief of these; it is a phenolic compound with antioxidant, anti-inflammatory, and anti-tumor properties.53 Gingerol suppresses inflammation by blocking the movement of NF-kB into the nucleus, with the resulting down-regulation of such inflammatory cytokines as TNF-alpha, as well as inducible nitric oxide synthase (iNOS).52,53 Another major ginger compound, zerumbone, suppresses NF-kB activation induced by a host of common carcinogens, blocking metastasis and invasion while increasing apoptosis.54
Ginger is a spice from southern Asia, that has many health-giving specific components.52 Gingerol is the chief of these; it is a phenolic compound with antioxidant, anti-inflammatory, and anti-tumor properties.53 Gingerol suppresses inflammation by blocking the movement of NF-kB into the nucleus, with the resulting down-regulation of such inflammatory cytokines as TNF-alpha, as well as inducible nitric oxide synthase (iNOS).52,53 Another major ginger compound, zerumbone, suppresses NF-kB activation induced by a host of common carcinogens, blocking metastasis and invasion while increasing apoptosis.54
Ginger extracts have been used to reduce the viability of gastric cancer cells, ovarian cancers, cancers of the breast, liver, colon, and lung, as well as UV light-induced skin cancers.55-60
 Here’s what we know in 2011 about the relationship between NF-kB and cancer, courtesy of Dr. Sahdeo Prasad of the Cytokine Research Laboratory at the MD Anderson Cancer Center in Houston:5
Here’s what we know in 2011 about the relationship between NF-kB and cancer, courtesy of Dr. Sahdeo Prasad of the Cytokine Research Laboratory at the MD Anderson Cancer Center in Houston:5
NF-kB becomes activated in response to a host of factors that account for as much as 95% of all cancers. These triggers include tobacco, stress, dietary components, obesity, alcohol, infections, radiation, and environmental toxins.1-3
NF-kB activation has been linked with the transformation of normal cells into cancer cells (thus NF-kB can be thought of as promoting cancer development).1,95
NF-kB is active at a higher rate in cancer cells than in healthy cells. A developing cancer also increases the amount of active NF-kB.6,13
NF-kB has been linked to the survival of dangerous cancer stem cells, the early “parent” cells of many cancers that are now known to exist in otherwise normal tissues. Cancer stem cells can “self-renew” more successfully than healthy cells.
NF-kB stimulates production of gene products that keep cancer cells from dying naturally through the process of programmed cell death, or apoptosis.96
NF-kB also increases production of gene products associated with proliferation, the rapid and repeated cell divisions that give cancers their aggressive growth characteristics.97
NF-kB controls expression of gene products linked with invasion, new blood vessel growth (angiogenesis), and metastatic spread of tumors.96,98-100
Many carcinogens activate NF-kB, but most effective cancer-preventing (chemopreventive) compounds suppress NF-kB activation.1,15,101
Prasad notes that these observations highlight just how intertwined NF-kB is with cancer growth and metastasis.5 Others have gone on to point out that, though a powerful nemesis, NF-kB also offers myriad opportunities to intervene powerfully and block cancers in their tracks.8,13 As we continue to clarify the close relationship between chronic inflammation and cancer, more and more such opportunities are becoming evident.93
Green Tea
 Green tea extracts are widely known for their benefits in preventing many common chronic diseases. Their chief constituents, the catechin family of polyphenols, are powerful antioxidants that are given credit for most of tea’s beneficial effects.61,62 Catechins act at many different targets.18,63,64 Inhibition of NF-kB by green tea catechins has recently been found to be a major mechanism by which they block each stage of carcinogenesis.63-65
Green tea extracts are widely known for their benefits in preventing many common chronic diseases. Their chief constituents, the catechin family of polyphenols, are powerful antioxidants that are given credit for most of tea’s beneficial effects.61,62 Catechins act at many different targets.18,63,64 Inhibition of NF-kB by green tea catechins has recently been found to be a major mechanism by which they block each stage of carcinogenesis.63-65
NF-kB inhibition by green tea extracts rich in epigallocatechin gallate (EGCG) has been of particular interest.866 EGCG has been shown to produce apoptosis in experimental prostate cancer, inhibit growth of squamous cell carcinomas of the head and neck, inhibit production and limit invasion of experimentally induced breast cancers, reduce the incidence of carcinogen-induced lung cancers, and sensitize melanoma cells to growth inhibition by other agents.67-71
Isoflavones
 Isoflavones and isoflavonoids are molecules found in many plants, particularly the bean family. Plants use them as disease-fighting compounds, and also as stimulants for the symbiotic bacteria that help them use nitrogen to build proteins. Isoflavones from soybeans have many beneficial and protective effects in humans as well. The low rate of hormone-related cancers (e.g., breast and prostate cancers) in Asian countries is partially attributed to the high consumption of soy isoflavones in most traditional Asian diets.72,73
Isoflavones and isoflavonoids are molecules found in many plants, particularly the bean family. Plants use them as disease-fighting compounds, and also as stimulants for the symbiotic bacteria that help them use nitrogen to build proteins. Isoflavones from soybeans have many beneficial and protective effects in humans as well. The low rate of hormone-related cancers (e.g., breast and prostate cancers) in Asian countries is partially attributed to the high consumption of soy isoflavones in most traditional Asian diets.72,73
Soy isoflavones like genistein are powerful modulators of NF-kB.73 Genistein acts both by preventing NF-kB’s movement into the nucleus to activate inflammatory genes and also by preventing earlier molecular events that would lead to its activation.74 Genistein promotes cancer cell death by apoptosis as a result of down-regulation of NF-kB.74
Isoflavone blockade of NF-kB effects contributes to prevention and modulation of colon75 and lung cancers.76 Isoflavone treatment also makes certain cancer cells increasingly susceptible to radiation and chemotherapy effects, largely because decreased NF-kB activity impairs the cancer cells’ survival signaling pathways.73,77,78
Omega-3 Fatty Acids
 Diets rich in omega-3 fatty acids from fish are associated with reduced risk of several types of cancer and other chronic conditions.79,80 The omega-3s are known to be powerful anti-inflammatory agents through their effects on the important inflammatory enzymes like COX-2 and 5-LOX.81 But recent research is also showing that omega-3s can inhibit NF-kB activation by multiple mechanisms, giving them still more powerful control over the inflammation that can lead to cancers.
Diets rich in omega-3 fatty acids from fish are associated with reduced risk of several types of cancer and other chronic conditions.79,80 The omega-3s are known to be powerful anti-inflammatory agents through their effects on the important inflammatory enzymes like COX-2 and 5-LOX.81 But recent research is also showing that omega-3s can inhibit NF-kB activation by multiple mechanisms, giving them still more powerful control over the inflammation that can lead to cancers.
Omega-3s are now associated with NF-kB-related reduction in inflammation-mediated growth of pancreatic, breast, prostate, colon, and skin cancer cells.82-90 In the case of pancreatic cancer, the omega-3 EPA preserved the integrity of the natural inhibitor of NF-kB.82 Omega-3s used in combination with conventional chemotherapy act synergistically to kill tumor cells, largely through NF-kB inhibition that promotes apoptosis.89,91
Summary
A breakthrough study published late last year revealed that inflammation is necessary in order for breast cancer cells to proliferate and metastasize. The master control complex called nuclear factor-kappaB or NF-kB triggers and regulates this inflammatory cascade, which has been implicated in the onset and development of 95% of all cancers.
NF-kB stimulates genes to produce inflammatory cytokines and other signaling molecules that promote cancer growth and development. Inhibition of NF-kB is a powerful target for cancer prevention and has been shown to reduce cancer-stimulative events at every level of carcinogenesis (cancer development). A broad array of natural dietary interventions powerfully inhibits pathologic NF-kB activity, comprising a critical strategy in the fight against cancer. Scores of new studies detail the effects of NF-kB inhibition on cancer prevention. The low rates of cancers in countries with high dietary intake of natural NF-kB inhibitors points strongly to the value of such agents as functional foods and as nutraceuticals.
In addition to the many natural products outlined here, there’s now strong evidence that vitamin D, Withania somnifera (ashwagandha), and pomegranate extracts have similar NF-kB-blocking capabilities.102-104
The good news for Life Extension members is that they have been using potent NF-kB-inhibiting nutrients for many decades.
Material used with permission of Life Extension. All rights reserved.
1. Shin SR, Sanchez-Velar N, Sherr DH, Sonenshein GE. 7,12-dimethylbenz(a)anthracene treatment of a c-rel mouse mammary tumor cell line induces epithelial to mesenchymal transition via activation of nuclear factor-kappaB. Cancer Res. 2006 Mar 1;66(5):2570-5.
2. Renehan AG, Roberts DL, Dive C. Obesity and cancer: pathophysiological and biological mechanisms. Arch Physiol Biochem. 2008 Feb;114(1):71-83.
3. Enwonwu CO, Meeks VI. Bionutrition and oral cancer in humans. Crit Rev Oral Biol Med. 1995;6(1):5-17.
4. Liu M, Sakamaki T, Casimiro MC, et al. The canonical NF-kappaB pathway governs mammary tumorigenesis in transgenic mice and tumor stem cell expansion. Cancer Res. 2010 Dec 15;70(24):10464-73.
5. Prasad S, Ravindran J, Aggarwal BB. NF-kappaB and cancer: how intimate is this relationship. Mol Cell Biochem. 2010 Mar;336(1-2):25-37.
6. Pianetti S, Arsura M, Romieu-Mourez R, Coffey RJ, Sonenshein GE. Her-2/neu overexpression induces NF-kappaB via a PI3-kinase/Akt pathway involving calpain-mediated degradation of IkappaB-alpha that can be inhibited by the tumor suppressor PTEN. Oncogene. 2001 Mar 15;20(11):1287-99.
7. Bhat-Nakshatri P, Sweeney CJ, Nakshatri H. Identification of signal transduction pathways involved in constitutive NF-kappaB activation in breast cancer cells. Oncogene. 2002 Mar 27;21(13):2066-78.
8. Ahmed KM, Cao N, Li JJ. HER-2 and NF-kappaB as the targets for therapy-resistant breast cancer. Anticancer Res. 2006 Nov-Dec;26(6B):4235-43.
9. Weldon CB, Burow ME, Rolfe KW, Clayton JL, Jaffe BM, Beckman BS. NF-kappa B-mediated chemoresistance in breast cancer cells. Surgery. 2001 Aug;130(2):143-50.
10. Plummer SM, Holloway KA, Manson MM, et al. Inhibition of cyclo-oxygenase 2 expression in colon cells by the chemopreventive agent curcumin involves inhibition of NF-kappaB activation via the NIK/IKK signalling complex. Oncogene. 1999 Oct 28;18(44):6013-20.
11. Dhanalakshmi S, Singh RP, Agarwal C, Agarwal R. Silibinin inhibits constitutive and TNFalpha-induced activation of NF-kappaB and sensitizes human prostate carcinoma DU145 cells to TNFalpha-induced apoptosis. Oncogene. 2002 Mar 7;21(11):1759-67.
12. Gukovsky I, Reyes CN, Vaquero EC, Gukovskaya AS, Pandol SJ. Curcumin ameliorates ethanol and nonethanol experimental pancreatitis. Am J Physiol Gastrointest Liver Physiol. 2003 Jan;284(1):G85-95.
13. Li SH, Hawthorne VS, Neal CL, et al. Upregulation of neutrophil gelatinase-associated lipocalin by ErbB2 through nuclear factor-kappaB activation. Cancer Res. 2009 Dec 15;69(24):9163-8.
14. Aravindaram K, Yang NS. Anti-inflammatory plant natural products for cancer therapy. Planta Med. 2010 Aug;76(11):1103-17.
15. Reuter S, Prasad S, Phromnoi K, et al. Thiocolchicoside exhibits anticancer effects through downregulation of NF-{kappa}B pathway and its regulated gene products linked to inflammation and cancer. Cancer Prev Res (Phila). 2010 Nov;3(11):1462-72.
16. Ralhan R, Pandey MK, Aggarwal BB. Nuclear factor-kappa B links carcinogenic and chemopreventive agents. Front Biosci (Schol Ed). 2009;1:45-60.
17. Aggarwal BB, Shishodia S. Suppression of the nuclear factor-kappaB activation pathway by spice-derived phytochemicals: reasoning for seasoning. Ann N Y Acad Sci. 2004 Dec;1030:434-41.
18. McCarty MF. Targeting multiple signaling pathways as a strategy for managing prostate cancer: multifocal signal modulation therapy. Integr Cancer Ther. 2004 Dec;3(4):349-80.
19. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. 2006 Mar 10;160(1):1-40.
20. Bouzyk E, Gradzka I, Iwanenko T, Kruszewski M, Sochanowicz B, Szumiel I. The response of L5178Y lymphoma sublines to oxidative stress: antioxidant defence, iron content and nuclear translocation of the p65 subunit of NF-kappaB. Acta Biochim Pol. 2000;47(4):881-8.
21. Saxena A, Saxena AK, Singh J, Bhushan S. Natural antioxidants synergistically enhance the anticancer potential of AP9-cd, a novel lignan composition from Cedrus deodara in human leukemia HL-60 cells. Chem Biol Interact. 2010 Dec 5;188(3):580-90.
22. Guo W, Kong E, Meydani M. Dietary polyphenols, inflammation, and cancer. Nutr Cancer. 2009 Nov;61(6):807-10.
23. Ho HH, Chang CS, Ho WC, Liao SY, Wu CH, Wang CJ. Anti-metastasis effects of gallic acid on gastric cancer cells involves inhibition of NF-kappaB activity and downregulation of PI3K/AKT/small GTPase signals. Food Chem Toxicol. 2010 Aug-Sep;48(8-9):2508-16.
24. Lin Y, Shi R, Wang X, Shen HM. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr Cancer Drug Targets. 2008 Nov;8(7):634-46.
25. Prasad AS, Beck FW, Snell DC, Kucuk O. Zinc in cancer prevention. Nutr Cancer. 2009 Nov;61(6):879-87.
26. Deep G, Gangar SC, Oberlies NH, Kroll DJ, Agarwal R. Isosilybin A induces apoptosis in human prostate cancer cells via targeting Akt, NF-kappaB, and androgen receptor signaling. Mol Carcinog. 2010 Oct;49(10):902-12.
27. Li L, Zeng J, Gao Y, He D. Targeting silibinin in the antiproliferative pathway. Expert Opin Investig Drugs. 2010 Feb;19(2):243-55.
28. Palozza P, Colangelo M, Simone R, et al. Lycopene induces cell growth inhibition by altering mevalonate pathway and Ras signaling in cancer cell lines. Carcinogenesis. 2010 Oct;31(10):1813-21.
29. Kunnumakkara AB, Sung B, Ravindran J, et al. {Gamma}-tocotrienol inhibits pancreatic tumors and sensitizes them to gemcitabine treatment by modulating the inflammatory microenvironment. Cancer Res. 2010 Nov 1;70(21):8695-705.
30. Shen T, Park YC, Kim SH, Lee J, Cho JY. Nuclear factor-kappaB/signal transducers and activators of transcription-1-mediated inflammatory responses in lipopolysaccharide-activated macrophages are a major inhibitory target of kahweol, a coffee diterpene. Biol Pharm Bull. 2010;33(7):1159-64.
31. Liu Y, Yadev VR, Aggarwal BB, Nair MG. Inhibitory effects of black pepper (Piper nigrum) extracts and compounds on human tumor cell proliferation, cyclooxygenase enzymes, lipid peroxidation and nuclear transcription factor-kappa-B. Nat Prod Commun. 2010 Aug;5(8):1253-7.
32. Duvoix A, Blasius R, Delhalle S, et al. Chemopreventive and therapeutic effects of curcumin. Cancer Lett. 2005 Jun 8;223(2):181-90.
33. Thangapazham RL, Sharma A, Maheshwari RK. Multiple molecular targets in cancer chemoprevention by curcumin. AAPS J. 2006;8(3):E443-9.
34. Kunnumakkara AB, Anand P, Aggarwal BB. Curcumin inhibits proliferation, invasion, angiogenesis and metastasis of different cancers through interaction with multiple cell signaling proteins. Cancer Lett. 2008 Oct 8;269(2):199-225.
35. Reuter S, Eifes S, Dicato M, Aggarwal BB, Diederich M. Modulation of anti-apoptotic and survival pathways by curcumin as a strategy to induce apoptosis in cancer cells. Biochem Pharmacol. 2008 Dec 1;76(11):1340-51.
36. Everett PC, Meyers JA, Makkinje A, Rabbi M, Lerner A. Preclinical assessment of curcumin as a potential therapy for B-CLL. Am J Hematol. 2007 Jan;82(1):23-30.
37. Dhillon N, Aggarwal BB, Newman RA, et al. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin Cancer Res. 2008 Jul 15;14(14):4491-9.
38. Kamat AM, Tharakan ST, Sung B, Aggarwal BB. Curcumin potentiates the antitumor effects of Bacillus Calmette-Guerin against bladder cancer through the downregulation of NF-kappaB and upregulation of TRAIL receptors. Cancer Res. 2009 Dec 1;69(23):8958-66.
39. Majumdar AP, Banerjee S, Nautiyal J, et al. Curcumin synergizes with resveratrol to inhibit colon cancer. Nutr Cancer. 2009;61(4):544-53.
40. Bisht S, Mizuma M, Feldmann G, et al. Systemic administration of polymeric nanoparticle-encapsulated curcumin (NanoCurc) blocks tumor growth and metastases in preclinical models of pancreatic cancer. Mol Cancer Ther. 2010 Aug;9(8):2255-64.
41. Jutooru I, Chadalapaka G, Lei P, Safe S. Inhibition of NFkappaB and pancreatic cancer cell and tumor growth by curcumin is dependent on specificity protein down-regulation. J Biol Chem. 2010 Aug 13;285(33):25332-44.
42. Ramachandran C, Resek AP, Escalon E, Aviram A, Melnick SJ. Potentiation of gemcitabine by Turmeric Force in pancreatic cancer cell lines. Oncol Rep. 2010 Jun;23(6):1529-35.
43. Suphim B, Prawan A, Kukongviriyapan U, Kongpetch S, Buranrat B, Kukongviriyapan V. Redox modulation and human bile duct cancer inhibition by curcumin. Food Chem Toxicol. 2010 Aug-Sep;48(8-9):2265-72.
44. Zhang C, Li B, Zhang X, Hazarika P, Aggarwal BB, Duvic M. Curcumin selectively induces apoptosis in cutaneous T-cell lymphoma cell lines and patients’ PBMCs: potential role for STAT-3 and NF-kappaB signaling. J Invest Dermatol. 2010 Aug;130(8):2110-9.
45. Aggarwal BB. Targeting inflammation-induced obesity and metabolic diseases by curcumin and other nutraceuticals. Annu Rev Nutr. 2010 Aug 21;30:173-99.
46. Borek C. Antioxidant health effects of aged garlic extract. J Nutr. 2001 Mar;131(3s):1010S-5S.
47. Ban JO, Lee HS, Jeong HS, et al. Thiacremonone augments chemotherapeutic agent-induced growth inhibition in human colon cancer cells through inactivation of nuclear factor-{kappa}B. Mol Cancer Res. 2009 Jun;7(6):870-9.
48. Pratheeshkumar P, Thejass P, Kutan G. Diallyl disulfide induces caspase-dependent apoptosis via mitochondria-mediated intrinsic pathway in B16F-10 melanoma cells by up-regulating p53, caspase-3 and down-regulating pro-inflammatory cytokines and nuclear factor-kappabeta-mediated Bcl-2 activation. J Environ Pathol Toxicol Oncol. 2010;29(2):113-25.
49. Ban JO, Yuk DY, Woo KS, et al. Inhibition of cell growth and induction of apoptosis via inactivation of NF-kappaB by a sulfurcompound isolated from garlic in human colon cancer cells. J Pharmacol Sci. 2007 Aug;104(4):374-83.
50. Karmakar S, Banik NL, Patel SJ, Ray SK. Garlic compounds induced calpain and intrinsic caspase cascade for apoptosis in human malignant neuroblastoma SH-SY5Y cells. Apoptosis. 2007 Apr;12(4):671-84.
51. Sriram N, Kalayarasan S, Ashokkumar P, Sureshkumar A, Sudhandiran G. Diallyl sulfide induces apoptosis in Colo 320 DM human colon cancer cells: involvement of caspase-3, NF-kappaB, and ERK-2. Mol Cell Biochem. 2008 Apr;311(1-2):157-65.
52. Kim SO, Kundu JK, Shin YK, et al. [6]-Gingerol inhibits COX-2 expression by blocking the activation of p38 MAP kinase and NF-kappaB in phorbol ester-stimulated mouse skin. Oncogene. 2005 Apr 7;24(15):2558-67.
53. Oyagbemi AA, Saba AB, Azeez OI. Molecular targets of [6]-gingerol: Its potential roles in cancer chemoprevention. Biofactors. 2010 May-Jun;36(3):169-78.
54. Takada Y, Murakami A, Aggarwal BB. Zerumbone abolishes NF-kappaB and IkappaBalpha kinase activation leading to suppression of antiapoptotic and metastatic gene expression, upregulation of apoptosis, and downregulation of invasion. Oncogene. 2005 Oct 20;24(46):6957-69.
55. Ishiguro K, Ando T, Maeda O, et al. Ginger ingredients reduce viability of gastric cancer cells via distinct mechanisms. Biochem Biophys Res Commun. 2007 Oct 12;362(1):218-23.
56. Kim JK, Kim Y, Na KM, Surh YJ, Kim TY. [6]-Gingerol prevents UVB-induced ROS production and COX-2 expression in vitro and in vivo. Free Radic Res. 2007 May;41(5):603-14.
57. Rhode J, Fogoros S, Zick S, et al. Ginger inhibits cell growth and modulates angiogenic factors in ovarian cancer cells. BMC Complement Altern Med. 2007;7:44.
58. Habib SH, Makpol S, Abdul Hamid NA, Das S, Ngah WZ, Yusof YA. Ginger extract (Zingiber officinale) has anti-cancer and anti-inflammatory effects on ethionine-induced hepatoma rats. Clinics (Sao Paulo). 2008 Dec;63(6):807-13.
59. Kim M, Miyamoto S, Yasui Y, Oyama T, Murakami A, Tanaka T. Zerumbone, a tropical ginger sesquiterpene, inhibits colon and lung carcinogenesis in mice. Int J Cancer. 2009 Jan 15;124(2):264-71.
60. Sung B, Murakami A, Oyajobi BO, Aggarwal BB. Zerumbone abolishes RANKL-induced NF-kappaB activation, inhibits osteoclastogenesis, and suppresses human breast cancer-induced bone loss in athymic nude mice. Cancer Res. 2009 Feb 15;69(4):1477-84.
61. Yang CS, Maliakal P, Meng X. Inhibition of carcinogenesis by tea. Annu Rev Pharmacol Toxicol. 2002;42:25-54.
62. Butt MS, Sultan MT. Green tea: nature’s defense against malignancies. Crit Rev Food Sci Nutr. 2009 May;49(5):463-73.
63. Khan N, Afaq F, Saleem M, Ahmad N, Mukhtar H. Targeting multiple signaling pathways by green tea polyphenol (-)-epigallocatechin-3-gallate. Cancer Res. 2006 Mar 1;66(5):2500-5.
64. Khan N, Mukhtar H. Multitargeted therapy of cancer by green tea polyphenols. Cancer Lett. 2008 Oct 8;269(2):269-80.
65. Navarro-Peran E, Cabezas-Herrera J, Sanchez-Del-Campo L, Garcia-Canovas F, Rodriguez-Lopez JN. The anti-inflammatory and anti-cancer properties of epigallocatechin-3-gallate are mediated by folate cycle disruption, adenosine release and NF-kappaB suppression. Inflamm Res. 2008 Oct;57(10):472-8.
66. Chen L, Zhang HY. Cancer preventive mechanisms of the green tea polyphenol (-)-epigallocatechin-3-gallate. Molecules. 2007;12(5):946-57.
67. Siddiqui IA, Shukla Y, Adhami VM, et al. Suppression of NFkappaB and its regulated gene products by oral administration of green tea polyphenols in an autochthonous mouse prostate cancer model. Pharm Res. 2008 Sep;25(9):2135-42.
68. Amin AR, Khuri FR, Chen ZG, Shin DM. Synergistic growth inhibition of squamous cell carcinoma of the head and neck by erlotinib and epigallocatechin-3-gallate: the role of p53-dependent inhibition of nuclear factor-kappaB. Cancer Prev Res (Phila). 2009 Jun;2(6):538-45.
69. Nihal M, Ahsan H, Siddiqui IA, Mukhtar H, Ahmad N, Wood GS. (-)-Epigallocatechin-3-gallate (EGCG) sensitizes melanoma cells to interferon induced growth inhibition in a mouse model of human melanoma. Cell Cycle. 2009 Jul 1;8(13):2057-63.
70. Sen T, Moulik S, Dutta A, et al. Multifunctional effect of epigallocatechin-3-gallate (EGCG) in downregulation of gelatinase-A (MMP-2) in human breast cancer cell line MCF-7. Life Sci. 2009 Feb 13;84(7-8):194-204.
71. Roy P, Nigam N, Singh M, et al. Tea polyphenols inhibit cyclooxygenase-2 expression and block activation of nuclear factor-kappa B and Akt in diethylnitrosoamine induced lung tumors in Swiss mice. Invest New Drugs. 2010 Aug;28(4):466-71.
72. Sarkar FH, Li Y. The role of isoflavones in cancer chemoprevention. Front Biosci. 2004 Sep 1;9:2714-24.
73. Banerjee S, Li Y, Wang Z, Sarkar FH. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008 Oct 8;269(2):226-42.
74. Li Z, Li J, Mo B, et al. Genistein induces cell apoptosis in MDA-MB-231 breast cancer cells via the mitogen-activated protein kinase pathway. Toxicol In Vitro. 2008 Oct;22(7):1749-53.
75. Ravasco P, Aranha MM, Borralho PM, et al. Colorectal cancer: can nutrients modulate NF-kappaB and apoptosis? Clin Nutr. 2010 Feb;29(1):42-6.
76. Gadgeel SM, Ali S, Philip PA, Wozniak A, Sarkar FH. Genistein enhances the effect of epidermal growth factor receptor tyrosine kinase inhibitors and inhibits nuclear factor kappa B in nonsmall cell lung cancer cell lines. Cancer. 2009 May 15;115(10):2165-76.
77. Singh-Gupta V, Zhang H, Banerjee S, et al. Radiation-induced HIF-1alpha cell survival pathway is inhibited by soy isoflavones in prostate cancer cells. Int J Cancer. 2009 Apr 1;124(7):1675-84.
78. Zhang B, Shi ZL, Liu B, Yan XB, Feng J, Tao HM. Enhanced anticancer effect of gemcitabine by genistein in osteosarcoma: the role of Akt and nuclear factor-kappaB. Anticancer Drugs. 2010 Mar;21(3):288-96.
79. Slagsvold JE, Pettersen CH, Storvold GL, Follestad T, Krokan HE, Schonberg SA. DHA alters expression of target proteins of cancer therapy in chemotherapy resistant SW620 colon cancer cells. Nutr Cancer. 2010 Jul;62(5):611-21.
80. Jho DH, Cole SM, Lee EM, Espat NJ. Role of omega-3 fatty acid supplementation in inflammation and malignancy. Integr Cancer Ther. 2004 Jun;3(2):98-111.
81. Groeger AL, Cipollina C, Cole MP, et al. Cyclooxygenase-2 generates anti-inflammatory mediators from omega-3 fatty acids. Nat Chem Biol. 2010 Jun;6(6):433-41.
82. Ross JA, Maingay JP, Fearon KC, Sangster K, Powell JJ. Eicosapentaenoic acid perturbs signalling via the NFkappaB transcriptional pathway in pancreatic tumour cells. Int J Oncol. 2003 Dec;23(6):1733-8.
83. Narayanan NK, Narayanan BA, Reddy BS. A combination of docosahexaenoic acid and celecoxib prevents prostate cancer cell growth in vitro and is associated with modulation of nuclear factor-kappaB, and steroid hormone receptors. Int J Oncol. 2005 Mar;26(3):785-92.
84. Schley PD, Jijon HB, Robinson LE, Field CJ. Mechanisms of omega-3 fatty acid-induced growth inhibition in MDA-MB-231 human breast cancer cells. Breast Cancer Res Treat. 2005 Jul;92(2):187-95.
85. Hering J, Garrean S, Dekoj TR, et al. Inhibition of proliferation by omega-3 fatty acids in chemoresistant pancreatic cancer cells. Ann Surg Oncol. 2007 Dec;14(12):3620-8.
86. Horia E, Watkins BA. Complementary actions of docosahexaenoic acid and genistein on COX-2, PGE2 and invasiveness in MDA-MB-231 breast cancer cells. Carcinogenesis. 2007 Apr;28(4):809-15.
87. Nowak J, Weylandt KH, Habbel P, et al. Colitis-associated colon tumorigenesis is suppressed in transgenic mice rich in endogenous n-3 fatty acids. Carcinogenesis. 2007 Sep;28(9):1991-5.
88. Kim HH, Lee Y, Eun HC, Chung JH. Eicosapentaenoic acid inhibits TNF-alpha-induced matrix metalloproteinase-9 expression in human keratinocytes, HaCaT cells. Biochem Biophys Res Commun. 2008 Apr 4;368(2):343-9.
89. Shaikh IA, Brown I, Schofield AC, Wahle KW, Heys SD. Docosahexaenoic acid enhances the efficacy of docetaxel in prostate cancer cells by modulation of apoptosis: the role of genes associated with the NF-kappaB pathway. Prostate. 2008 Nov 1;68(15):1635-46.
90. Ghosh-Choudhury T, Mandal CC, Woodruff K, et al. Fish oil targets PTEN to regulate NFkappaB for downregulation of anti-apoptotic genes in breast tumor growth. Breast Cancer Res Treat. 2009 Nov;118(1):213-28.
91. Ding WQ, Liu B, Vaught JL, Palmiter RD, Lind SE. Clioquinol and docosahexaenoic acid act synergistically to kill tumor cells. Mol Cancer Ther. 2006 Jul;5(7):1864-72.
92. Dong G, Chen Z, Kato T, Van Waes C. The host environment promotes the constitutive activation of nuclear factor-kappaB and proinflammatory cytokine expression during metastatic tumor progression of murine squamous cell carcinoma. Cancer Res. 1999 Jul 15;59(14):3495-504.
93. Pikarsky E, Porat RM, Stein I, et al. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature. 2004 Sep 23;431(7007):461-6.
94. Chapkin RS, Kim W, Lupton JR, McMurray DN. Dietary docosahexaenoic and eicosapentaenoic acid: emerging mediators of inflammation. Prostaglandins Leukot Essent Fatty Acids. 2009 Aug-Sep;81(2-3):187-91.
95. Pratt MA, Tibbo E, Robertson SJ, et al. The canonical NF-kappaB pathway is required for formation of luminal mammary neoplasias and is activated in the mammary progenitor population. Oncogene. 2009 Jul 30;28(30):2710-22.
96. deGraffenried LA, Chandrasekar B, Friedrichs WE, et al. NF-kappa B inhibition markedly enhances sensitivity of resistant breast cancer tumor cells to tamoxifen. Ann Oncol. 2004 Jun;15(6):885-90.
97. Brantley DM, Chen CL, Muraoka RS, et al. Nuclear factor-kappaB (NF-kappaB) regulates proliferation and branching in mouse mammary epithelium. Mol Biol Cell. 2001 May;12(5):1445-55.
98. Hagemann T, Wilson J, Kulbe H, et al. Macrophages induce invasiveness of epithelial cancer cells via NF-kappa B and JNK. J Immunol. 2005 Jul 15;175(2):1197-205.
99. Liu M, Ju X, Willmarth NE, et al. Nuclear factor-kappaB enhances ErbB2-induced mammary tumorigenesis and neoangiogenesis in vivo. Am J Pathol. 2009 May;174(5):1910-20.
100. Beshir AB, Ren G, Magpusao AN, Barone LM, Yeung KC, Fenteany G. Raf kinase inhibitor protein suppresses nuclear factor-kappaB-dependent cancer cell invasion through negative regulation of matrix metalloproteinase expression. Cancer Lett. 2010 Dec 28;299(2):137-49.
101. Tapia MA, Gonzalez-Navarrete I, Dalmases A, et al. Inhibition of the canonical IKK/NF kappa B pathway sensitizes human cancer cells to doxorubicin. Cell Cycle. 2007 Sep 15;6(18):2284-92.
102. Mulabagal V, Subbaraju GV, Rao CV, et al. Withanolide sulfoxide from Aswagandha roots inhibits nuclear transcription factor-kappa-B, cyclooxygenase and tumor cell proliferation. Phytother Res. 2009 Jul;23(7):987-92.
103. Kwon HJ, Won YS, Suh HW, et al. Vitamin D3 upregulated protein 1 suppresses TNF-alpha-induced NF-kappaB activation in hepatocarcinogenesis. J Immunol. 2010 Oct 1;185(7):3980-9.
104. Rettig MB, Heber D, An J, et al. Pomegranate extract inhibits androgen-independent prostate cancer growth through a nuclear factor-kappaB-dependent mechanism. Mol Cancer Ther. 2008 Sep;7(9):2662-71.
Related products
-

Super Absorbable Tocotrienols
Price: 30,25 €Support healthy hair growth & more Derived from red palm oil Provide... -

Pomegranate Complete
Price: 29,28 €Standardized extracts from the whole fruit, flower and seed oil... -

Mega Green Tea Extract (decaffeinated)
Price: 31,23 €One capsule provides more polyphenols than seven cups of green tea... -

Zinc Caps
Price: 14,13 €Support your body's natural immune defenses Supports body’s natural immune... -

Curcumin Elite™ Turmeric Extract
Price: 35,14 €This proprietary curcumin formula is the best we’ve ever offered Free... -

Super Omega-3 EPA/DHA with Sesame Lignans &...
Price: 43,64 €Get enough omega-3s Powerful cardiovascular and cognitive health support... -

Advanced Curcumin Elite™ Turmeric Extract,...
Price: 26,10 €Highly absorbable curcumin with complementary plant extracts Offers 45x... -

Optimized Ashwagandha Extract
Price: 12,83 €Enhances mental energy and concentration Is believed to confer...
Related posts
-
 Rebuild Aging Joints!
In this article, you will learn about UC-II®, or undenatured type II collagen, a novel intervention for both...
Rebuild Aging Joints!
In this article, you will learn about UC-II®, or undenatured type II collagen, a novel intervention for both... -
Is Conventional Pomegranate Extract Enough?
An abundance of clinical data has established pomegranate as a broad-spectrum, multi-modal defense against the...
-
Next-Generation Nutrients for Rapid Relief of Joint Pain
In recent years, adults seeking relief from the crippling pain of arthritis have had to choose between expensive,...
-
Lipoic Acid Reverses Mitochondrial Decay
It is estimated that 85% of the oxygen contained in every breath you take is consumed by the mitochondria within each...
-
Vitamin C and Dihydroquercetin
Every day, our bodies are under continual assault by damaging agents known as free radicals. Generally, both internal...